The Social Brain Paradigm
- First Online: 20 November 2019

Cite this chapter

- Sal Restivo 2
597 Accesses
This chapter introduces my perspective on and my model of the social brain. The development of the social brain paradigm reflects a general development from hierarchical to network thinking across the intellectual spectrum during the latter part of the twentieth century. I discuss the evolution of the social intelligence hypothesis into the social brain hypothesis, and the reigning myths about the brain that have obstructed social brain thinking. I review the key developments in the history of neuroscience at its nexus with the life- and social sciences and their connections to social brain research and theory. The chapter ends with a presentation of my model of the social brain as a networked information system situated in and coupled with a social ecology (Appendix 1 ). In Appendix 2 , I review the concept of connectomics, and in Appendix 3 , I list links to glossaries on brain terminology to aid readers in understanding the terms used in the text to describe the structure and function of the brain.
- Social intelligence
- Social brain
- Connectomics
- Information
This is a preview of subscription content, log in via an institution to check access.
Access this chapter
- Available as PDF
- Read on any device
- Instant download
- Own it forever
- Available as EPUB and PDF
- Compact, lightweight edition
- Dispatched in 3 to 5 business days
- Free shipping worldwide - see info
- Durable hardcover edition
Tax calculation will be finalised at checkout
Purchases are for personal use only
Institutional subscriptions
Barta, R. 2014. Anthropology of the Brain: Consciousness, Culture, and Free Will . Cambridge: Cambridge University Press.
Book Google Scholar
Brothers, L. 1990. The Social Brain: A Project for Integrating Primate Behavior and Neurophysiology in a New Domain. Concepts in Neuroscience I: 27–51.
Google Scholar
Brüne, M., H. Ribbert, and W. Schiefenhövel, eds. 2003. The Social Brain: Evolution and Pathology . West Sussex: John Wiley & Sons.
Cacioppo, J.T., G.G. Berntson, R. Adolphs, et al., eds. 2002. Foundations in Social Neuroscience . Cambridge, MA: MIT Press.
Carhart-Harris, R.L., and J. Friston. 2019. REBUS and the Anarchic Brain: Toward a Unified Model of the Brain Action of Psychedelics. Pharmacological Reviews 71 (3): 316–344.
Article Google Scholar
Dean, S. 2018. The Human Brain Can Create Structures in Up to 11 Dimensions. https://www.sciencealert.com/science-discovers-human-brain-works-up-to-11-dimensions .
Duke, D.W., and W.S. Pritchard, eds. 1991. Measuring Chaos in the Brain . London: World Scientific.
Dunbar, R., C. Gamble, and J.G. Owlett, eds. 2010. Social Brain: Distributed Mind . Oxford: Oxford University Press.
Ferrari, P.F., and G. Rizzolatti. 2014. Mirror Neuron Research: The Past and the Future. Philosophical Transactions of the Royal Society B 369 (1644). https://doi.org/10.1098/rstb.2013.0169 .
———. 2015. New Frontiers in Mirror Neurons Research . Oxford: Oxford University Press.
Feynman, R. 1979. The Smartest Man in the World . OMNI interview; reprinted as pp. 189–204 in Richard Feynman (1999). The Pleasure of Finding Things Out . New York: Basic Books.
Fiske and Taylor. 2013. Social Cognition from Brains to Culture . London: Sage.
Franks, D.D. and T. Smith, eds. (1999), Mind, Brain, and Society: Towards a Neurosociology of Emotion, Vol. 5 of Social Perspectives on Emotion (Stamford, CT: JAI Press).
Franks, D.D., and J.H. Turner, eds. 2013. Handbook of Neurosociology . New York: Springer.
Geertz, C. 1973. The Interpretation of Cultures . New York: Basic Books.
———. 2000. Available Light . Princeton: Princeton University Press.
Hagmann, P. 2005. From Diffusion MRI to Brain Connectomics Hampshire . PhD Thesis. Lausanne: Ecole Polytechnique Fédérale de Lausanne.
Hyeonlin, J., and L. Seung-Hwan. 2018. From Neurons to Social Beings: Short Review of the Mirror Neuron System Research and its Socio-Psychological and Psychiatric Implications. Clinical Psychopharmacological Neuroscience 16 (1): 18–31.
Jacob, F. 1977. Evolution and Tinkering. Science 196 (4295): 1161–1166.
Johnson-Ulrich, L. 2017. The Social Intelligence Hypothesis. In Encyclopedia of Evolutionary Psychological Science , ed. T.K. Shackelford and V.A. Weekes-Shackelford, 1–7. New York: Springer.
Kull, K. 1998. On Semiosis, Umwelt, and Semiosphere. Semiotica 120 (3/4): 299–310.
Khanna, P. 2016. Connectography: Mapping the Future of Global Civilization . New York: Random House.
Lehnertz, K., C.E. Elger, J. Arnhold, and P. Grassberger, eds. 2000. Chaos in Brain? Proceedings of the Workshop . University of Bonn: Germany.
Markram, H. 2008. Fixing the Location and Dimensions of Functional Neocortical Columns. Human Frontiers Science Program Journal 2 (3): 132–135.
Pickersgill, M., and I.V. Keulen, eds. 2012. Sociological Reflections on the Neurosciences , Advances in Medical Sociology. Vol. 13. Bingley, UK: Emerald Group Publishing, Ltd.
Racine, V. 2014. Evolution and tinkering (1977), by Francois Jacob. Embryo Project Encyclopedia. https://embryo.asu.edu/pages/evolution-and-tinkering-1977-francois-jacob .
Reimann, M.W., M. Nolte, et al. 2017. Cliques of Neurons Bound into Cavities Provide a Missing Link Between Structure and Function. Frontiers in Computational Neuroscience 11 (48): 1–16.
Restivo, S., S. Weiss, and A. Stingl. 2014. Worlds of ScienceCraft . New York: Routledge.
Schutt, R.K., L. Seidman, and M.S. Keshavan, eds. 2015. Social Neuroscience: Brain, Mind, and Society . Cambridge, MA: Harvard University Press.
Sebeok, Thomas A. 1976. Foreword. In Contributions to the Doctrine of Signs , ed. T. Sebeok, x. Lanham, NH: University Press of America.
Sebeok, Thomas A., and D.J. Umlker-Sebeok. 1978. Linguistics: Speech Surrogates: Drum and Whistle Systems. American Anthropologist 80 (3): 712–713.
Seung, S. 2012. Connectome: How the Brain’s Wiring Makes Us Who We Are . New York: Houghton Mifflin Harcourt.
Skarda, C.A., and W.J. Freeman. 1987. How Brains Make Chaos in Order to Make Sense of the World. Behavioral and Brain Sciences 10: 161–195.
———. 1990. Chaos and the New Science of the Brain. Concepts in Neuroscience 2: 275–285.
Soresi, E. 2014. The Anarchic Brain . Milano: Bookrepublic.
Sporns, O., G. Tononi, and R. Kötter. 2005. The Human Connectome: A Structural Description of the Human Brain. PLoS Computational Biology 1 (4): e42.
Taylor, J.M. 2016. Mirror Neurons After a Quarter Century: New Light, New Cracks. http://sitn.hms.harvard.edu/flash/2016/mirror-neurons-quarter-century-new-light-new-cracks/ .
von Uexküll, Thure. 1987. The Sign Theory of Jakob von Uexküll. In Classics of Semiotics , ed. M. Krampen et al., 147–179. New York: Plenum.
Chapter Google Scholar
———. 1957. A Stroll Through the Worlds of Animals and Men: A Picture Book of Invisible Worlds. In Instinctive Behavior: The Development of a Modern Concept , ed. Claire H. Schiller, 5–80. New York: International Universities Press.
von Uexküll, J. 2010/1934. A Foray into the World of Animals and Humans With a Theory of Meaning . Trans. J.D. O’Neil. Minneapolis: University of Minnesota Press.
de Waal, P.L. Tyack, et al., eds. 2003. Animal Social Complexity: Intelligence, Culture and Individualized Societies . Cambridge, MA: Harvard University Press.
Whiten, A. 2000: 185–196; discussion 196–201. Social Complexity and Social Intelligence . Basel, Switzerland: Novartis Foundation Symposium 233.
Zapporoli, L., M. Porta, and E. Paulesu. 2015. The Anarchic Brain in Action: The Contribution of Task-Based fMRI Studies to the Understanding of Gilles de la Tourette Syndrome. Current Opinion in Neurology 6: 604–611.
Download references
Author information
Authors and affiliations.
Ridgewood, NY, USA
Sal Restivo
You can also search for this author in PubMed Google Scholar
Appendix 1: The Social Brain Model: The Social Ecology of the Brain

The networked social brain
Figure 5.1 The original version of this model was designed with Sabrina Weiss . I have taken it through a number of revisions designed to keep pace with developments in neuroscience and in social neuroscience and neurosociology. It was designed to graphically represent and expand Clifford Geertz’s argument for the synchronic emergence of brain and culture. In review, then: (1) biological, social, and cultural causal forces are reciprocally intertwined and conjointly causal; (2) human behavioral repertoires emerge from the complex parallel and recursive interactions of cells, genes, neurons, neural nets, organs, biomes, the brain and central nervous system, other elements of the body’s systems and subsystems, and our social interactions in their ecological and umwelt contexts; (3) socialization is re-imagined as a process that simultaneously informs and variably integrates the biological self, the neurological self, and the social self to construct personality and character; (4) each element in the model is a dialectical entity containing its own internal “seeds” of change, and as following a temporal dynamic that may be at different times synchronous or dyssynchronous relative to other elements; (5) each element is conceived as an information system with all systems multiply inter-linked by the circulation of information; (6) the diagonals with double-headed arrows which crisscross the model map the chaotic dynamics and cooperative neural mass discussed by C.A. Skarda and W.J. Freeman ( 1987 , 1990 ); (7) the unit model is activated in a triad of unit models and it is that triad that is the basic model of brain/mind/culture/world. This reflects the idea that the triad is the basic unit of social life (Restivo et al. 2014 : 104n1); and (8) the diagram is the General Connectome . A connectome maps the elements and interconnections in a network. The term has been used specifically in connection with mapping the neural connections in the brain. Connectome s may range in scale from maps of parts of the nervous system to a map of all of the neural interactions in the brain. Partial connectome s have been constructed of the retina and primary visual cortex of the mouse. In line with these developments, my model represents the highest level of the connectome , a connectome of connectome s.
Based on the ideas introduced in the previous chapters I can now offer an initial concept formula for the probability of an “innovative thought.” iT p = qc 2 × K + G, where qc 2 is the amount of cultural capital the person commands and K is a constant that represents the cultural context and network structure the person is embedded in; qc 2 because doubling the amount of cultural capital, for example, quadruples its impact factor. K = C + N t . C = Cultural Context, an index that takes into account a variety of demographic, class, gender, and institutional diversity indicators; N = the density and diversity of the network structure of the society. G = the genius cluster quotient at time t. When considering the etiology of behaviors traditionally considered to be genetically grounded, it is now important to recognize that the brain, like humans, arrives on the evolutionary stage always, already, and everywhere, social. Therefore, what we have considered to be linearly transmitted genetic phenomena must now be viewed in the context of a brain that is at no stage of development separated from the social and cultural imperatives that form us. The very notions of “genes” and “genetic” must now be revised in the context of the social brain paradigm.
The next stage in this project is to embed the basic triad of the General Connectome in the nested networks of the social and cultural connectomes locally, regionally, and globally so that we now visualize a Global Connectome driven by the circulation of information across nested networks. On the rationale for a global connectome (my interpretation), see Khanna ( 2016 ) on “connectography.”
Appendix 2: Connectomics
A connectome comprehensively maps neural connections in the brain. More broadly, a connectome maps all the neural connections in an organism’s nervous system. Hagmann ( 2005 ) and Sporns et al. ( 2005 ) independently and simultaneously introduced the term “connectome ,” inspired by the efforts to construct a genome. Connectomics is the science of assembling and analyzing connectome data sets. Hagmann and Sporns discussed research strategies for developing comprehensive structural descriptions of the brain’s networks, a dataset they called the “connectome .” Such a connectome would help us understand the emergence of functional brain states from their structural substrate. Connectomic s, the production and study of connectome s, can be applied at different scales from the full set of neurons and synapses in a part or all of an organism’s nervous system to macro-level descriptions of the connections between all cortical and subcortical structures. The full connectome of the roundworm has been constructed along with partial connectome s of a mouse retina and primary visual cortex.
Appendix 3: Guides to the Technical Brain Terms Used in this Book
https://www.dana.org/brainglossary/
https://www.brainfacts.org/glossary
https://mayfieldclinic.com/pe-anatbrain.htm
https://www.google.com/search?q=parts+of+the+brain+and+their+functions+chart&safe=off&client=safari&sa=X&rls=en&biw=1218&bih=752&tbm=isch&source=iu&ictx=1&fir=nYBnlzaBiedEcM%253A%252CdE9NmhvLUJFzjM%252C_&vet=1&usg=AI4_-kTdU8tqEE14iJcfb1jdKW-J-0HWcg&ved=2ahUKEwiU3s709YTjAhUMZd8KHR-TDJcQ9QEwBXoECAcQDg#imgrc=_&vet=1
Appendix 4: Bibliographic Notes for Chapter 5
The social intelligence hypothesis.
Websites: for an overview of the social intelligence hypothesis and some initial references to evolution and the social brain see: https://www.sciencedirect.com/topics/psychology/social-intelligence-hypothesis ; https://www.sciencedirect.com/science/article/pii/S1364661306003263 : more recent examinations of the social intelligence hypothesis suggest the need for a broader theoretical framework that embraces “both inter-specific differences and similarities in cognition; … how selection pressures that are associated with sociality interact with those that are imposed by non-social forms of environmental complexity, and how both types of functional demands interact with phylogenetic and developmental constraints” (Holekamp: https://doi.org/10.1016/j.tics.2006.11.003 ). For a critical evaluation of the hypothesis by Hemelrijk (2007) see: https://pdfs.semanticscholar.org/8df0/5c96641806c2bc45b220aeb792f563ee6473.pdf
Books, Articles, and Chapters
Whiten ( 2000 ), de Waal et al. ( 2003 ), Johnson-Ulrich ( 2017 ).
On the meaning of “monotonic”: In mathematics, a monotonic function (or monotone function) is a function between ordered sets that preserves or reverses the given order. In calculus, a function defined as a subset of the real numbers with real values is called monotonic if and only if it is either entirely non-increasing or entirely non-decreasing. That is, a function that increases monotonically does not exclusively have to increase, it simply must not decrease.
The Social Brain Paradigm: Selected References
The literature on social cognition is relevant here but tends traditionally to be too embedded in the psychological-biological-neuroscience context which trumps the social and cultural contextual approaches. For an important exception, see Fiske and Taylor ( 2013 ); the authors are still under the influence of the idea that biology has causal priority over culture; but this is a serious effort to integrate emerging developments in social cognition with developments in social neuroscience, cultural psychology, and applied psychology. On the social brain per se, see Brüne et al. ( 2003 ), Dunbar et al. ( 2010 ); on social neuroscience, see Cacioppo et al. ( 2002 ), Schutt et al. ( 2015 ); on neurosociology, see Franks and Smith ( 1999 ), Franks and Turner ( 2013 ), Pickersgill and Keulen ( 2012 ), Barta ( 2014 ): this is really an anthropology of consciousness and extremely well done except for Bartra’s effort to save free will. On Issues Surrounding the Idea of the Split Brain
https://www.health.harvard.edu/blog/right-brainleft-brain-right- ; https://brainconnection.brainhq.com/2001/06/26/roger-sperry-the-brains-inside-the-brain
https://www.inc.com/jessica-stillman/left-brained-v-right-brained-people-is-a-total-myt.html The Chaotic and Anarchic Brain Skarda and Freeman ( 1987 , 1990 ), Duke and Pritchard ( 1991 ), Lehnertz et al. ( 2000 ), Soresi ( 2014 ), Zapporoli et al. ( 2015 ), Carhart-Harris and Friston ( 2019 ).
On the Concept of the Umwelt
See: Kull ( 1998 ), Sebeok ( 1976 ), Sebeok and Umlker-Sebeok ( 1978 ), and Uexküll, J.v. ( 1957 , 2010/1934 ), and Uexküll, T.v. ( 1987 ).
Rights and permissions
Reprints and permissions
Copyright information
© 2020 The Author(s)
About this chapter
Restivo, S. (2020). The Social Brain Paradigm. In: Einstein’s Brain . Palgrave Pivot, Cham. https://doi.org/10.1007/978-3-030-32918-1_5
Download citation
DOI : https://doi.org/10.1007/978-3-030-32918-1_5
Published : 20 November 2019
Publisher Name : Palgrave Pivot, Cham
Print ISBN : 978-3-030-32917-4
Online ISBN : 978-3-030-32918-1
eBook Packages : Behavioral Science and Psychology Behavioral Science and Psychology (R0)
Share this chapter
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
- Publish with us
Policies and ethics
- Find a journal
- Track your research
The Social Brain Hypothesis and Human Evolution
Primate societies are unusually complex compared to those of other animals, and the need to manage such complexity is the main explanation for the fact that primates have unusually large brains. Primate sociality is based on bonded relationships that underpin coalitions, which in turn are designed to buffer individuals against the social stresses of living in large, stable groups. This is reflected in a correlation between social group size and neocortex size in primates (but not other species of animals), commonly known as the social brain hypothesis, although this relationship itself is the outcome of an underlying relationship between brain size and behavioral complexity. The relationship between brain size and group size is mediated, in humans at least, by mentalizing skills. Neuropsychologically, these are all associated with the size of units within the theory of mind network (linking prefrontal cortex and temporal lobe units). In addition, primate sociality involves a dual-process mechanism whereby the endorphin system provides a psychopharmacological platform off which the cognitive component is then built. This article considers the implications of these findings for the evolution of human cognition over the course of hominin evolution.
- Related Documents
Social Brain, Distributed Mind
To understand who we are and why we are, we need to understand both modern humans and the ancestral stages that brought us to this point. The core to that story has been the role of evolving cognition — the social brain — in mediating the changes in behaviour that we see in the archaeological record. This volume brings together two powerful approaches — the social brain hypothesis and the concept of the distributed mind. The volume compares perspectives on these two approaches from a range of disciplines, including archaeology, psychology, philosophy, sociology and the cognitive and evolutionary sciences. A particular focus is on the role that material culture plays as a scaffold for distributed cognition, and how almost three million years of artefact and tool use provides the data for tracing key changes in areas such as language, technology, kinship, music, social networks and the politics of local, everyday interaction in small-world societies. A second focus is on how, during the course of hominin evolution, increasingly large spatially distributed communities created stresses that threatened social cohesion. This volume offers the possibility of new insights into the evolution of human cognition and social lives that will further our understanding of the relationship between mind and world.
Is the Social Brain Theory Applicable to Human Individual Differences? Relationship between Sociability Personality Dimension and Brain Size
Assessing sources of error in comparative analyses of primate behavior: intraspecific variation in group size and the social brain hypothesis, the social brain.
The first discussion of a relationship between sociality and intelligence came in the middle of the twentieth century, especially by Humphrey who suggested that living socially demanded intellectual abilities above and beyond those required by an animal’s ecology. This led to the Social Intelligence Hypothesis, and then the Machiavellian Intelligence Hypothesis, both proposing that sociality was the main driver of the superior intellect of primates, especially humans. Two key challenges for this hypothesis are that sociality is difficult to quantify and cognition is not well tested by problem solving. More importantly, as data from more species have been examined, the analyses increasingly fail to show that sociality explains variation in brain size, even in primates. I conclude that appealing as this hypothesis is, it does not do a very compelling job of explaining variation in brain size.
Theory of mind in autism, schizophrenia, and in-between
AbstractAutism and schizophrenia are presented as the extremes of disorders affecting the social brain. By viewing human cognition impairment in terms of competence and performance, a variety of social brain disorders can be identified along the autistic-psychotic continuum.
Understanding primate brain evolution
We present a detailed reanalysis of the comparative brain data for primates, and develop a model using path analysis that seeks to present the coevolution of primate brain (neocortex) and sociality within a broader ecological and life-history framework. We show that body size, basal metabolic rate and life history act as constraints on brain evolution and through this influence the coevolution of neocortex size and group size. However, they do not determine either of these variables, which appear to be locked in a tight coevolutionary system. We show that, within primates, this relationship is specific to the neocortex. Nonetheless, there are important constraints on brain evolution; we use path analysis to show that, in order to evolve a large neocortex, a species must first evolve a large brain to support that neocortex and this in turn requires adjustments in diet (to provide the energy needed) and life history (to allow sufficient time both for brain growth and for ‘software’ programming). We review a wider literature demonstrating a tight coevolutionary relationship between brain size and sociality in a range of mammalian taxa, but emphasize that the social brain hypothesis is not about the relationship between brain/neocortex size and group size per se ; rather, it is about social complexity and we adduce evidence to support this. Finally, we consider the wider issue of how mammalian (and primate) brains evolve in order to localize the social effects.
Coevolution of cultural intelligence, extended life history, sociality, and brain size in primates
Explanations for primate brain expansion and the evolution of human cognition and culture remain contentious despite extensive research. While multiple comparative analyses have investigated variation in brain size across primate species, very few have addressed why primates vary in how much they use social learning. Here, we evaluate the hypothesis that the enhanced reliance on socially transmitted behavior observed in some primates has coevolved with enlarged brains, complex sociality, and extended lifespans. Using recently developed phylogenetic comparative methods we show that, across primate species, a measure of social learning proclivity increases with absolute and relative brain volume, longevity (specifically reproductive lifespan), and social group size, correcting for research effort. We also confirm relationships of absolute and relative brain volume with longevity (both juvenile period and reproductive lifespan) and social group size, although longevity is generally the stronger predictor. Relationships between social learning, brain volume, and longevity remain when controlling for maternal investment and are therefore not simply explained as a by-product of the generally slower life history expected for larger brained species. Our findings suggest that both brain expansion and high reliance on culturally transmitted behavior coevolved with sociality and extended lifespan in primates. This coevolution is consistent with the hypothesis that the evolution of large brains, sociality, and long lifespans has promoted reliance on culture, with reliance on culture in turn driving further increases in brain volume, cognitive abilities, and lifespans in some primate lineages.
Why Humans Aren’t Just Great Apes
Although we share many aspects of our behaviour and biology with our primate cousins, humans are, nonetheless, different in one crucial respect: our capacity to live in the world of the imagination. This is reflected in two core aspects of our behaviour that are in many ways archetypal of what it is to be human: religion and story-telling. I shall show how these remarkable traits seem to have arisen as a natural development of the social brain hypothesis, and the underlying nature of primate sociality and cognition, as human societies have been forced to expand in size during the course of our evolution over the past 5 million years.
When Individuals Do Not Stop at the Skin
This chapter examines contemporary hunter-gatherer societies in Africa and elsewhere in light of the social brain and the distributed mind hypotheses. One question asked is whether African hunter-gatherers offer the best model for societies at the dawn of symbolic culture, or whether societies elsewhere offer better models. The chapter argues for the former. Theoretical concepts touched on include sharing and exchange, universal kin classification, and the relation between group size and social networks. The chapter offers reinterpretations of classic anthropological notions such as Wissler's age-area hypothesis, Durkheim's collective consciousness and Lévi-Strauss's elementary structures of kinship. Finally, the chapter outlines a theory of the co-evolution of language and kinship through three phases (signifying, syntactic and symbolic) and the subsequent breakdown of the principles of the symbolic phase across much of the globe in Neolithic times.
Absolute, not relative brain size correlates with sociality in ground squirrels
The social brain hypothesis (SBH) contends that cognitive demands associated with living in cohesive social groups favour the evolution of large brains. Although the correlation between relative brain size and sociality reported in various groups of birds and mammals provides broad empirical support for this hypothesis, it has never been tested in rodents, the largest mammalian order. Here, we test the predictions of the SBH in the ground squirrels from the tribe Marmotini. These rodents exhibit levels of sociality ranging from solitary and single-family female kin groups to egalitarian polygynous harems but feature similar ecologies and life-history traits. We found little support for the association between increase in sociality and increase in relative brain size. Thus, sociality does not drive the evolution of encephalization in this group of rodents, a finding inconsistent with the SBH. However, body mass and absolute brain size increase with sociality. These findings suggest that increased social complexity in the ground squirrels goes hand in hand with larger body mass and brain size, which are tightly coupled to each other.
Export Citation Format
Share document.
The social brain hypothesis and its implications for social evolution
Affiliation.
- 1 Institute of Cognitive & Evolutionary Anthropology, University of Oxford, Oxford, UK. [email protected]
- PMID: 19575315
- DOI: 10.1080/03014460902960289
The social brain hypothesis was proposed as an explanation for the fact that primates have unusually large brains for body size compared to all other vertebrates: Primates evolved large brains to manage their unusually complex social systems. Although this proposal has been generalized to all vertebrate taxa as an explanation for brain evolution, recent analyses suggest that the social brain hypothesis takes a very different form in other mammals and birds than it does in anthropoid primates. In primates, there is a quantitative relationship between brain size and social group size (group size is a monotonic function of brain size), presumably because the cognitive demands of sociality place a constraint on the number of individuals that can be maintained in a coherent group. In other mammals and birds, the relationship is a qualitative one: Large brains are associated with categorical differences in mating system, with species that have pairbonded mating systems having the largest brains. It seems that anthropoid primates may have generalized the bonding processes that characterize monogamous pairbonds to other non-reproductive relationships ('friendships'), thereby giving rise to the quantitative relationship between group size and brain size that we find in this taxon. This raises issues about why bonded relationships are cognitively so demanding (and, indeed, raises questions about what a bonded relationship actually is), and when and why primates undertook this change in social style.
Publication types
- Biological Evolution*
- Brain / physiology*
- Primates / physiology*
- Primates / psychology
- Social Behavior*
Thank you for visiting nature.com. You are using a browser version with limited support for CSS. To obtain the best experience, we recommend you use a more up to date browser (or turn off compatibility mode in Internet Explorer). In the meantime, to ensure continued support, we are displaying the site without styles and JavaScript.
- View all journals
- Explore content
- About the journal
- Publish with us
- Sign up for alerts
- Perspective
- Published: 27 October 2023
How deep is the brain? The shallow brain hypothesis
- Mototaka Suzuki ORCID: orcid.org/0000-0002-2151-4882 1 ,
- Cyriel M. A. Pennartz ORCID: orcid.org/0000-0001-8328-1175 1 &
- Jaan Aru ORCID: orcid.org/0000-0003-3927-452X 2
Nature Reviews Neuroscience volume 24 , pages 778–791 ( 2023 ) Cite this article
17k Accesses
2 Citations
261 Altmetric
Metrics details
- Computational neuroscience
- Neural circuits
- Neurophysiology
- Sensorimotor processing
Deep learning and predictive coding architectures commonly assume that inference in neural networks is hierarchical. However, largely neglected in deep learning and predictive coding architectures is the neurobiological evidence that all hierarchical cortical areas, higher or lower, project to and receive signals directly from subcortical areas. Given these neuroanatomical facts, today’s dominance of cortico-centric, hierarchical architectures in deep learning and predictive coding networks is highly questionable; such architectures are likely to be missing essential computational principles the brain uses. In this Perspective, we present the shallow brain hypothesis: hierarchical cortical processing is integrated with a massively parallel process to which subcortical areas substantially contribute. This shallow architecture exploits the computational capacity of cortical microcircuits and thalamo-cortical loops that are not included in typical hierarchical deep learning and predictive coding networks. We argue that the shallow brain architecture provides several critical benefits over deep hierarchical structures and a more complete depiction of how mammalian brains achieve fast and flexible computational capabilities.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
24,99 € / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
176,64 € per year
only 14,72 € per issue
Buy this article
Purchase on Springer Link
Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout
Similar content being viewed by others

If deep learning is the answer, what is the question?
Andrew Saxe, Stephanie Nelli & Christopher Summerfield

Brain-optimized deep neural network models of human visual areas learn non-hierarchical representations
Ghislain St-Yves, Emily J. Allen, … Thomas Naselaris

A deep learning framework for neuroscience
Blake A. Richards, Timothy P. Lillicrap, … Konrad P. Kording
Hegde, J. & Felleman, D. J. Reappraising the functional implications of the primate visual anatomical hierarchy. Neuroscientist 13 , 416–421 (2007).
Article PubMed Google Scholar
LeCun, Y., Bengio, Y. & Hinton, G. Deep learning. Nature 521 , 436–444 (2015).
Article CAS PubMed Google Scholar
Stokel-Walker, C. & Van Noorden, R. What ChatGPT and generative AI mean for science. Nature 614 , 214–216 (2023).
He, K., Zhang, X., Ren, S. & Sun, J. Deep residual learning for image recognition. In 2016 IEEE Conference on Computer Vision and Pattern Recognition (CVPR) 770–778 (2016).
Russakovsky, O. et al. ImageNet large scale visual recognition challenge. Int. J. Comput. Vis. 115 , 211–252 (2015).
Article Google Scholar
Xu, K. et al. Show, attend and tell: neural image caption generation with visual attention. In 32nd Int. Conf. on Machine Learning (eds F. Bach. & D. Blei) 2048–2057 (2015).
Fukushima, K. Neocognitron—a self-organizing neural network model for a mechanism of pattern-recognition unaffected by shift in position. Biol. Cybern. 36 , 193–202 (1980).
Hubel, D. H. & Wiesel, T. N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J. Physiol. 160 , 106–154 (1962).
Article CAS PubMed PubMed Central Google Scholar
Chang, L. & Tsao, D. Y. The code for facial identity in the primate brain. Cell 169 , 1013–1028.e14 (2017).
Yamins, D. L. et al. Performance-optimized hierarchical models predict neural responses in higher visual cortex. Proc. Natl Acad. Sci. USA 111 , 8619–8624 (2014).
Guclu, U. & van Gerven, M. A. Deep neural networks reveal a gradient in the complexity of neural representations across the ventral stream. J. Neurosci. 35 , 10005–10014 (2015).
Article PubMed PubMed Central Google Scholar
Cichy, R. M., Khosla, A., Pantazis, D., Torralba, A. & Oliva, A. Comparison of deep neural networks to spatio-temporal cortical dynamics of human visual object recognition reveals hierarchical correspondence. Sci. Rep. 6 , 27755 (2016).
Schrimpf, M. et al. The neural architecture of language: integrative modeling converges on predictive processing. Proc. Natl Acad. Sci. USA 118 , e2015646118 (2021).
Kriegeskorte, N. Deep neural networks: a new framework for modeling biological vision and brain information processing. Annu. Rev. Vis. Sci. 1 , 417–446 (2015).
Yamins, D. L. & DiCarlo, J. J. Using goal-driven deep learning models to understand sensory cortex. Nat. Neurosci. 19 , 356–365 (2016).
Friston, K. A theory of cortical responses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 360 , 815–836 (2005).
Rao, R. P. & Ballard, D. H. Predictive coding in the visual cortex: a functional interpretation of some extra-classical receptive-field effects. Nat. Neurosci. 2 , 79–87 (1999).
Lee, T. S. & Mumford, D. Hierarchical Bayesian inference in the visual cortex. J. Opt. Soc. Am. A Opt. Image Sci. Vis. 20 , 1434–1448 (2003).
Srinivasan, M. V., Laughlin, S. B. & Dubs, A. Predictive coding: a fresh view of inhibition in the retina. Proc. R. Soc. Lond. B Biol. Sci. 216 , 427–459 (1982).
Dayan, P., Hinton, G. E., Neal, R. M. & Zemel, R. S. The Helmholtz machine. Neural Comput. 7 , 889–904 (1995).
Dora, S., Bohte, S. M. & Pennartz, C. M. A. Deep gated Hebbian predictive coding accounts for emergence of complex neural response properties along the visual cortical hierarchy. Front. Comput. Neurosci. 15 , 666131 (2021).
McDermott, J. H., Wrobleski, D. & Oxenham, A. J. Recovering sound sources from embedded repetition. Proc. Natl Acad. Sci. USA 108 , 1188–1193 (2011).
Mill, R. W., Bohm, T. M., Bendixen, A., Winkler, I. & Denham, S. L. Modelling the emergence and dynamics of perceptual organisation in auditory streaming. PLoS Comput. Biol. 9 , e1002925 (2013).
Kanai, R., Komura, Y., Shipp, S. & Friston, K. Cerebral hierarchies: predictive processing, precision and the pulvinar. Philos. Trans. R. Soc. Lond. B Biol. Sci. 370 , 20140169 (2015).
Schwartenbeck, P., FitzGerald, T. H., Mathys, C., Dolan, R. & Friston, K. The dopaminergic midbrain encodes the expected certainty about desired outcomes. Cereb. Cortex 25 , 3434–3445 (2015).
Rikhye, R. V., Wimmer, R. D. & Halassa, M. M. Toward an integrative theory of thalamic function. Annu. Rev. Neurosci. 41 , 163–183 (2018).
Felleman, D. J. & Van Essen, D. C. Distributed hierarchical processing in the primate cerebral cortex. Cereb. Cortex 1 , 1–47 (1991).
Minsky, M. & Papert, S. Perceptrons; An Introduction to Computational Geometry (MIT Press, 1969).
Gross, C. G., Rocha-Miranda, C. E. & Bender, D. B. Visual properties of neurons in inferotemporal cortex of the macaque. J. Neurophysiol. 35 , 96–111 (1972).
Tsao, D. Y., Schweers, N., Moeller, S. & Freiwald, W. A. Patches of face-selective cortex in the macaque frontal lobe. Nat. Neurosci. 11 , 877–879 (2008).
Hegde, J. & Van Essen, D. C. A comparative study of shape representation in macaque visual areas V2 and V4. Cereb. Cortex 17 , 1100–1116 (2007).
Rockland, K. S. & Pandya, D. N. Laminar origins and terminations of cortical connections of the occipital lobe in the rhesus monkey. Brain Res. 179 , 3–20 (1979).
Markov, N. T. et al. Cortical high-density counterstream architectures. Science 342 , 1238406 (2013).
Markov, N. T. & Kennedy, H. The importance of being hierarchical. Curr. Opin. Neurobiol. 23 , 187–194 (2013).
D’Souza, R. D. et al. Hierarchical and nonhierarchical features of the mouse visual cortical network. Nat. Commun. 13 , 503 (2022).
Siegle, J. H. et al. Survey of spiking in the mouse visual system reveals functional hierarchy. Nature 592 , 86–92 (2021).
Nakamura, H., Gattass, R., Desimone, R. & Ungerleider, L. G. The modular organization of projections from areas V1 and V2 to areas V4 and TEO in macaques. J. Neurosci. 13 , 3681–3691 (1993).
Burkhalter, A., D’Souza, R. D., Ji, W. & Meier, A. M. Integration of feedforward and feedback information streams in the modular architecture of mouse visual cortex. Annu. Rev. Neurosci. 46 , 259–280 (2023).
Coogan, T. A. & Burkhalter, A. Hierarchical organization of areas in rat visual cortex. J. Neurosci. 13 , 3749–3772 (1993).
Friston, K. The free-energy principle: a unified brain theory? Nat. Rev. Neurosci. 11 , 127–138 (2010).
Bastos, A. M. et al. Canonical microcircuits for predictive coding. Neuron 76 , 695–711 (2012).
Pennartz, C. M. A., Dora, S., Muckli, L. & Lorteije, J. A. M. Towards a unified view on pathways and functions of neural recurrent processing. Trends Neurosci. 42 , 589–603 (2019).
Findling, C. et al. Brain-wide representations of prior information in mouse decision-making. Preprint at bioRxiv https://doi.org/10.1101/2023.07.04.547684 (2023).
Keller, G. B. & Mrsic-Flogel, T. D. Predictive processing: a canonical cortical computation. Neuron 100 , 424–435 (2018).
Keller, G. B., Bonhoeffer, T. & Hubener, M. Sensorimotor mismatch signals in primary visual cortex of the behaving mouse. Neuron 74 , 809–815 (2012).
Jordan, R. & Keller, G. B. Opposing influence of top-down and bottom-up input on excitatory layer 2/3 neurons in mouse primary visual cortex. Neuron 108 , 1194–1206.e5 (2020).
Padamsey, Z. & Rochefort, N. L. Defying expectations: how neurons compute prediction errors in visual cortex. Neuron 108 , 1016–1019 (2020).
Muzzu, T. & Saleem, A. B. Feature selectivity can explain mismatch signals in mouse visual cortex. Cell Rep. 37 , 109772 (2021).
Walsh, K. S., McGovern, D. P., Clark, A. & O’Connell, R. G. Evaluating the neurophysiological evidence for predictive processing as a model of perception. Ann. N. Y. Acad. Sci. 1464 , 242–268 (2020).
Schwiedrzik, C. M. & Freiwald, W. A. High-level prediction signals in a low-level area of the macaque face-processing hierarchy. Neuron 96 , 89–97.e4 (2017).
Issa, E. B., Cadieu, C. F. & DiCarlo, J. J. Neural dynamics at successive stages of the ventral visual stream are consistent with hierarchical error signals. eLife 7 , e42870 (2018).
Chao, Z. C., Takaura, K., Wang, L., Fujii, N. & Dehaene, S. Large-scale cortical networks for hierarchical prediction and prediction error in the primate brain. Neuron 100 , 1252–1266.e3 (2018).
Spratling, M. W. A review of predictive coding algorithms. Brain Cogn. 112 , 92–97 (2017).
Spratling, M. W. Fitting predictive coding to the neurophysiological data. Brain Res. 1720 , 146313 (2019).
Bianchini, M. & Scarselli, F. On the complexity of neural network classifiers: a comparison between shallow and deep architectures. IEEE Trans. Neural Netw. Learn. Syst. 25 , 1553–1565 (2014).
Cohen, N., Sharir, O. & Shashua, A. On the expressive power of deep learning: a tensor analysis. In 29th Annual Conference on Learning Theory (eds. Feldman, V., Rakhlin, A. & Shamir, O.) 698–728 (2016).
Hinton, G. E. Training products of experts by minimizing contrastive divergence. Neural Comput. 14 , 1771–1800 (2002).
Dabelow, L. & Ueda, M. Three learning stages and accuracy-efficiency tradeoff of restricted Boltzmann machines. Nat. Commun. 13 , 5474 (2022).
Liao, R., Kornblith, S., Ren, M., Fleet, D. J. & Hinton, G. Gaussian–Bernoulli RBMs without tears. Preprint at arXiv https://doi.org/10.48550/ARXIV.2210.10318 (2022).
Hilgetag, C. C. & Goulas, A. ‘Hierarchy’ in the organization of brain networks. Philos. Trans. R. Soc. Lond. B Biol. Sci. 375 , 20190319 (2020).
Sherman, S. M. & Guillery, R. W. Functional organization of thalamocortical relays. J. Neurophysiol. 76 , 1367–1395 (1996).
Jones, E. G. The thalamic matrix and thalamocortical synchrony. Trends Neurosci. 24 , 595–601 (2001).
Sherman, S. M. & Guillery, R. W. Exploring the Thalamus and Its Role in Cortical Function 2nd edn (MIT Press, 2009).
Halassa, M. Thalamus 1st edn (Cambridge Univ. Press, 2023).
Kemp, J. M. & Powell, T. P. The cortico-striate projection in the monkey. Brain 93 , 525–546 (1970).
Oka, H. Organization of the cortico-caudate projections. A horseradish peroxidase study in the cat. Exp. Brain Res. 40 , 203–208 (1980).
Ito, S. & Feldheim, D. A. The mouse superior colliculus: an emerging model for studying circuit formation and function. Front. Neural Circuits 12 , 10 (2018).
Basso, M. A. & May, P. J. Circuits for action and cognition: a view from the superior colliculus. Annu. Rev. Vis. Sci. 3 , 197–226 (2017).
May, P. J. The mammalian superior colliculus: laminar structure and connections. Prog. Brain Res. 151 , 321–378 (2006).
McBride, E. G. et al. Influence of claustrum on cortex varies by area, layer, and cell type. Neuron 111 , 275–290.e5 (2022).
Narikiyo, K. et al. The claustrum coordinates cortical slow-wave activity. Nat. Neurosci. 23 , 741–753 (2020).
Jackson, J., Karnani, M. M., Zemelman, B. V., Burdakov, D. & Lee, A. K. Inhibitory control of prefrontal cortex by the claustrum. Neuron 99 , 1029–1039.e4 (2018).
Legg, C. R., Mercier, B. & Glickstein, M. Corticopontine projection in the rat: the distribution of labelled cortical cells after large injections of horseradish peroxidase in the pontine nuclei. J. Comp. Neurol. 286 , 427–441 (1989).
Habas, C. & Cabanis, E. A. Cortical projections to the human red nucleus: a diffusion tensor tractography study with a 1.5-T MRI machine. Neuroradiology 48 , 755–762 (2006).
Tervo, D. G. et al. A designer AAV variant permits efficient retrograde access to projection neurons. Neuron 92 , 372–382 (2016).
Murakami, T., Matsui, T., Uemura, M. & Ohki, K. Modular strategy for development of the hierarchical visual network in mice. Nature 608 , 578–585 (2022).
Tang, L. & Higley, M. J. Layer 5 circuits in V1 differentially control visuomotor behavior. Neuron 105 , 346–354.e5 (2020).
Takahashi, N. et al. Active dendritic currents gate descending cortical outputs in perception. Nat. Neurosci. 23 , 1277–1285 (2020).
Fuster, J. M. The Prefrontal Cortex: Anatomy, Physiology, and Neuropsychology of the Frontal Lobe (Raven, 1980).
Miller, E. K. & Cohen, J. D. An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 24 , 167–202 (2001).
Oswald, M. J., Tantirigama, M. L., Sonntag, I., Hughes, S. M. & Empson, R. M. Diversity of layer 5 projection neurons in the mouse motor cortex. Front. Cell Neurosci. 7 , 174 (2013).
Akintunde, A. & Buxton, D. F. Origins and collateralization of corticospinal, corticopontine, corticorubral and corticostriatal tracts: a multiple retrograde fluorescent tracing study. Brain Res. 586 , 208–218 (1992).
Harris, K. D. & Shepherd, G. M. The neocortical circuit: themes and variations. Nat. Neurosci. 18 , 170–181 (2015).
Musall, S. et al. Pyramidal cell types drive functionally distinct cortical activity patterns during decision-making. Nat. Neurosci. 26 , 495–505 (2023).
CAS PubMed PubMed Central Google Scholar
Mohan, H. et al. Cortical glutamatergic projection neuron types contribute to distinct functional subnetworks. Nat. Neurosci. 26 , 481–494 (2023).
Kuramoto, E. et al. Ventral medial nucleus neurons send thalamocortical afferents more widely and more preferentially to layer 1 than neurons of the ventral anterior–ventral lateral nuclear complex in the rat. Cereb. Cortex 25 , 221–235 (2015).
Cruikshank, S. J. et al. Thalamic control of layer 1 circuits in prefrontal cortex. J. Neurosci. 32 , 17813–17823 (2012).
Schroeder, A. et al. Inhibitory top-down projections from zona incerta mediate neocortical memory. Neuron 111 , 727–738.e8 (2023).
Ahmadlou, M. et al. A cell type-specific cortico-subcortical brain circuit for investigatory and novelty-seeking behavior. Science 372 , eabe9681 (2021).
Brenner, J. M., Beltramo, R., Gerfen, C. R., Ruediger, S. & Scanziani, M. A genetically defined tecto-thalamic pathway drives a system of superior-colliculus-dependent visual cortices. Neuron 111 , 2247–2257.e7 (2023).
Guo, Z. V. et al. Maintenance of persistent activity in a frontal thalamocortical loop. Nature 545 , 181–186 (2017).
Hsiao, K. et al. A thalamic orphan receptor drives variability in short-term memory. Cell 183 , 522–536.e19 (2020).
Redinbaugh, M. J. et al. Thalamus modulates consciousness via layer-specific control of cortex. Neuron 106 , 66–75.e12 (2020).
Aru, J., Suzuki, M. & Larkum, M. E. Cellular mechanisms of conscious processing. Trends Cogn. Sci. 24 , 814–825 (2020).
Aru, J., Suzuki, M., Rutiku, R., Larkum, M. E. & Bachmann, T. Coupling the state and contents of consciousness. Front. Syst. Neurosci. 13 , 43 (2019).
Suzuki, M. & Larkum, M. E. General anesthesia decouples cortical pyramidal neurons. Cell 180 , 666–676.e13 (2020).
Schiff, N. D. et al. Behavioural improvements with thalamic stimulation after severe traumatic brain injury. Nature 448 , 600–603 (2007).
Bastos, A. M. et al. Neural effects of propofol-induced unconsciousness and its reversal using thalamic stimulation. eLife 10 , e60824 (2021).
Crick, F. C. & Koch, C. What is the function of the claustrum. Philos. Trans. R. Soc. Lond. B Biol. Sci. 360 , 1271–1279 (2005).
Chevee, M., Finkel, E. A., Kim, S. J., O’Connor, D. H. & Brown, S. P. Neural activity in the mouse claustrum in a cross-modal sensory selection task. Neuron 110 , 486–501.e7 (2022).
Huang, W., Qin, J., Zhang, C., Qin, H. & Xie, P. Footshock-induced activation of the claustrum–entorhinal cortical pathway in freely moving mice. Physiol. Res. 71 , 695–701 (2022).
Smythies, J. On the function of object cells in the claustrum—key components in information processing in the visual system? Front. Cell Neurosci. 9 , 443 (2015).
Tsumoto, T. & Suda, K. Effects of stimulation of the dorsocaudal claustrum on activities of striate cortex neurons in the cat. Brain Res. 240 , 345–349 (1982).
Remedios, R., Logothetis, N. K. & Kayser, C. A role of the claustrum in auditory scene analysis by reflecting sensory change. Front. Syst. Neurosci. 8 , 44 (2014).
Qadir, H. et al. The mouse claustrum synaptically connects cortical network motifs. Cell Rep. 41 , 111860 (2022).
Taylor, N. L. et al. Structural connections between the noradrenergic and cholinergic system shape the dynamics of functional brain networks. Neuroimage 260 , 119455 (2022).
Deutch, A. Y. & Roth, R. H. in Fundamental Neuroscience (eds M. J. Zigmond et al.) 193–234 (Academic, 1999).
Chevalier, G. & Deniau, J. M. Disinhibition as a basic process in the expression of striatal functions. Trends Neurosci. 13 , 277–280 (1990).
Voorn, P., Vanderschuren, L. J., Groenewegen, H. J., Robbins, T. W. & Pennartz, C. M. Putting a spin on the dorsal–ventral divide of the striatum. Trends Neurosci. 27 , 468–474 (2004).
Budinger, E., Heil, P., Hess, A. & Scheich, H. Multisensory processing via early cortical stages: connections of the primary auditory cortical field with other sensory systems. Neuroscience 143 , 1065–1083 (2006).
Benavidez, N. L. et al. Organization of the inputs and outputs of the mouse superior colliculus. Nat. Commun. 12 , 4004 (2021).
Beltramo, R. & Scanziani, M. A collicular visual cortex: neocortical space for an ancient midbrain visual structure. Science 363 , 64–69 (2019).
Constantinople, C. M. & Bruno, R. M. Effects and mechanisms of wakefulness on local cortical networks. Neuron 69 , 1061–1068 (2011).
Aru, J., Siclari, F., Phillips, W. A. & Storm, J. F. Apical drive—a cellular mechanism of dreaming? Neurosci. Biobehav. Rev. 119 , 440–455 (2020).
Wainstein, G., Muller, E. J., Taylor, N., Munn, B. & Shine, J. M. The role of the locus coeruleus in shaping adaptive cortical melodies. Trends Cogn. Sci. 26 , 527–538 (2022).
Polack, P. O., Friedman, J. & Golshani, P. Cellular mechanisms of brain state-dependent gain modulation in visual cortex. Nat. Neurosci. 16 , 1331–1339 (2013).
Harris, K. D. & Thiele, A. Cortical state and attention. Nat. Rev. Neurosci. 12 , 509–523 (2011).
Parikh, V., Kozak, R., Martinez, V. & Sarter, M. Prefrontal acetylcholine release controls cue detection on multiple timescales. Neuron 56 , 141–154 (2007).
Puig, M. V. & Gulledge, A. T. Serotonin and prefrontal cortex function: neurons, networks, and circuits. Mol. Neurobiol. 44 , 449–464 (2011).
Buhot, M. C., Martin, S. & Segu, L. Role of serotonin in memory impairment. Ann. Med. 32 , 210–221 (2000).
Petroni, F., Panzeri, S., Hilgetag, C. C., Kotter, R. & Young, M. P. Simultaneity of responses in a hierarchical visual network. Neuroreport 12 , 2753–2759 (2001).
Zeki, S. The rough seas of cortical cartography. Trends Neurosci. 41 , 242–244 (2018).
Silvanto, J. Why is “blindsight” blind? A new perspective on primary visual cortex, recurrent activity and visual awareness. Conscious. Cogn. 32 , 15–32 (2015).
Schmolesky, M. T. et al. Signal timing across the macaque visual system. J. Neurophysiol. 79 , 3272–3278 (1998).
Bullier, J. & Nowak, L. G. Parallel versus serial processing: new vistas on the distributed organization of the visual system. Curr. Opin. Neurobiol. 5 , 497–503 (1995).
Douglas, R. J. & Martin, K. A. Mapping the matrix: the ways of neocortex. Neuron 56 , 226–238 (2007).
Rockland, K. S. & Ichinohe, N. Some thoughts on cortical minicolumns. Exp. Brain Res. 158 , 265–277 (2004).
Molnár, Z. & Rockland, K. S. in Neural Circuit and Cognitive Development Ch. 5 (eds J. Rubenstein, P. Rakic, B. Chen & K. Y. Kwan) 103–126 (Academic, 2020).
Trojanowski, J. Q. & Jacobson, S. Medial pulvinar afferents to frontal eye fields in rhesus monkey demonstrated by horseradish peroxidase. Brain Res. 80 , 395–411 (1974).
Baizer, J. S., Desimone, R. & Ungerleider, L. G. Comparison of subcortical connections of inferior temporal and posterior parietal cortex in monkeys. Vis. Neurosci. 10 , 59–72 (1993).
Stanton, G. B., Goldberg, M. E. & Bruce, C. J. Frontal eye field efferents in the macaque monkey: I. Subcortical pathways and topography of striatal and thalamic terminal fields. J. Comp. Neurol. 271 , 473–492 (1988).
Lynch, J. C., Hoover, J. E. & Strick, P. L. Input to the primate frontal eye field from the substantia nigra, superior colliculus, and dentate nucleus demonstrated by transneuronal transport. Exp. Brain Res. 100 , 181–186 (1994).
Berman, R. A. & Wurtz, R. H. Exploring the pulvinar path to visual cortex. Prog. Brain Res. 171 , 467–473 (2008).
Huerta, M. F., Krubitzer, L. A. & Kaas, J. H. Frontal eye field as defined by intracortical microstimulation in squirrel monkeys, owl monkeys, and macaque monkeys: I. Subcortical connections. J. Comp. Neurol. 253 , 415–439 (1986).
Leichnetz, G. R., Smith, D. J. & Spencer, R. F. Cortical projections to the paramedian tegmental and basilar pons in the monkey. J. Comp. Neurol. 228 , 388–408 (1984).
Andersen, R. A., Asanuma, C., Essick, G. & Siegel, R. M. Corticocortical connections of anatomically and physiologically defined subdivisions within the inferior parietal lobule. J. Comp. Neurol. 296 , 65–113 (1990).
Lynch, J. C., Graybiel, A. M. & Lobeck, L. J. The differential projection of two cytoarchitectonic subregions of the inferior parietal lobule of macaque upon the deep layers of the superior colliculus. J. Comp. Neurol. 235 , 241–254 (1985).
Schall, J. D., Morel, A., King, D. J. & Bullier, J. Topography of visual cortex connections with frontal eye field in macaque: convergence and segregation of processing streams. J. Neurosci. 15 , 4464–4487 (1995).
Vernet, M., Quentin, R., Chanes, L., Mitsumasu, A. & Valero-Cabre, A. Frontal eye field, where art thou? Anatomy, function, and non-invasive manipulation of frontal regions involved in eye movements and associated cognitive operations. Front. Integr. Neurosci. 8 , 66 (2014).
PubMed PubMed Central Google Scholar
Liu, Y., Yttri, E. A. & Snyder, L. H. Intention and attention: different functional roles for LIPd and LIPv. Nat. Neurosci. 13 , 495–500 (2010).
Coe, B. C. & Munoz, D. P. Mechanisms of saccade suppression revealed in the anti-saccade task. Philos. Trans. R. Soc. Lond. B Biol. Sci. 372 , 20160192 (2017).
Milardi, D. et al. Red nucleus connectivity as revealed by constrained spherical deconvolution tractography. Neurosci. Lett. 626 , 68–73 (2016).
Na, J., Kakei, S. & Shinoda, Y. Cerebellar input to corticothalamic neurons in layers V and VI in the motor cortex. Neurosci. Res. 28 , 77–91 (1997).
Martinez-Gonzalez, C., Bolam, J. P. & Mena-Segovia, J. Topographical organization of the pedunculopontine nucleus. Front. Neuroanat. 5 , 22 (2011).
Sherman, S. M. & Guillery, R. W. Distinct functions for direct and transthalamic corticocortical connections. J. Neurophysiol. 106 , 1068–1077 (2011).
de Kock, C. P., Bruno, R. M., Spors, H. & Sakmann, B. Layer- and cell-type-specific suprathreshold stimulus representation in rat primary somatosensory cortex. J. Physiol. 581 , 139–154 (2007).
Masamizu, Y. et al. Two distinct layer-specific dynamics of cortical ensembles during learning of a motor task. Nat. Neurosci. 17 , 987–994 (2014).
Guo, K., Yamawaki, N., Svoboda, K. & Shepherd, G. M. G. Anterolateral motor cortex connects with a medial subdivision of ventromedial thalamus through cell type-specific circuits, forming an excitatory thalamo-cortico-thalamic loop via layer 1 apical tuft dendrites of layer 5b pyramidal tract type neurons. J. Neurosci. 38 , 8787–8797 (2018).
Bharioke, A. et al. General anesthesia globally synchronizes activity selectively in layer 5 cortical pyramidal neurons. Neuron 110 , 2024–2040.e10 (2022).
Larkum, M. A cellular mechanism for cortical associations: an organizing principle for the cerebral cortex. Trends Neurosci. 36 , 141–151 (2013).
Brea, J., Gaal, A. T., Urbanczik, R. & Senn, W. Prospective coding by spiking neurons. PLoS Comput. Biol. 12 , e1005003 (2016).
Roelfsema, P. R. & Holtmaat, A. Control of synaptic plasticity in deep cortical networks. Nat. Rev. Neurosci. 19 , 166–180 (2018).
Lillicrap, T. P., Santoro, A., Marris, L., Akerman, C. J. & Hinton, G. Backpropagation and the brain. Nat. Rev. Neurosci. 21 , 335–346 (2020).
Whittington, J. C. R. & Bogacz, R. Theories of error back-propagation in the brain. Trends Cogn. Sci. 23 , 235–250 (2019).
Xiong, Q., Znamenskiy, P. & Zador, A. M. Selective corticostriatal plasticity during acquisition of an auditory discrimination task. Nature 521 , 348–351 (2015).
Cox, J. & Witten, I. B. Striatal circuits for reward learning and decision-making. Nat. Rev. Neurosci. 20 , 482–494 (2019).
Park, J. M. et al. Deep and superficial layers of the primary somatosensory cortex are critical for whisker-based texture discrimination in mice. Preprint at bioRxiv https://doi.org/10.1101/2020.08.12.245381 (2022).
Hong, Y. K., Lacefield, C. O., Rodgers, C. C. & Bruno, R. M. Sensation, movement and learning in the absence of barrel cortex. Nature 561 , 542–546 (2018).
Von Neumann, J. The Computer and the Brain (Yale Univ. Press, 1958).
Mo, C. & Sherman, S. M. A sensorimotor pathway via higher-order thalamus. J. Neurosci. 39 , 692–704 (2019).
Lake, B. M., Ullman, T. D., Tenenbaum, J. B. & Gershman, S. J. Building machines that learn and think like people. Behav. Brain Sci. 40 , e253 (2017).
Baroni, M. Linguistic generalization and compositionality in modern artificial neural networks. Philos. Trans. R. Soc. Lond. B Biol. Sci. 375 , 20190307 (2020).
Ruediger, S. & Scanziani, M. Learning speed and detection sensitivity controlled by distinct cortico-fugal neurons in visual cortex. eLife 9 , e59247 (2020).
Brooks, R. A. A robust layered control-system for a mobile robot. IEEE T Robotic Autom. 2 , 14–23 (1986).
Brooks, R. A. New approaches to robotics. Science 253 , 1227–1232 (1991).
Haider, P., Ellenberger, B., Kriener, L., Jordan, J., Senn, W. & Petrovici, M. A. Latent equilibrium: a unified learning theory for arbitrarily fast computation with arbitrarily slow neurons. Adv. Neural Inf. Process. Syst. 34 , 17839–17851 (2021).
Google Scholar
Narayanan, R. T. et al. Beyond columnar organization: cell type- and target layer-specific principles of horizontal axon projection patterns in rat vibrissal cortex. Cereb. Cortex 25 , 4450–4468 (2015).
Chen, G., Scherr, F. & Maass, W. A data-based large-scale model for primary visual cortex enables brain-like robust and versatile visual processing. Sci. Adv. 8 , eabq7592 (2022).
Guest, J. M., Bast, A., Narayanan, R. T. & Oberlaender, M. Thalamus gates active dendritic computations in cortex during sensory processing. Preprint at bioRxiv https://doi.org/10.1101/2021.10.21.465325 (2021).
Constantinople, C. M. & Bruno, R. M. Deep cortical layers are activated directly by thalamus. Science 340 , 1591–1594 (2013).
Pluta, S. et al. A direct translaminar inhibitory circuit tunes cortical output. Nat. Neurosci. 18 , 1631–1640 (2015).
Stuart, G., Spruston, N. & Häusser, M. Dendrites 3rd edn (Oxford Univ. Press, 2016).
Major, G., Larkum, M. E. & Schiller, J. Active properties of neocortical pyramidal neuron dendrites. Annu. Rev. Neurosci. 36 , 1–24 (2013).
Mikulasch, F. A., Rudelt, L., Wibral, M. & Priesemann, V. Where is the error? Hierarchical predictive coding through dendritic error computation. Trends Neurosci. 46 , 45–59 (2022).
Richards, B. A. & Lillicrap, T. P. Dendritic solutions to the credit assignment problem. Curr. Opin. Neurobiol. 54 , 28–36 (2019).
Guerguiev, J., Lillicrap, T. P. & Richards, B. A. Towards deep learning with segregated dendrites. eLife 6 , e22901 (2017).
Hawkins, J. & Ahmad, S. Why neurons have thousands of synapses, a theory of sequence memory in neocortex. Front. Neural Circuits 10 , 23 (2016).
Schiess, M., Urbanczik, R. & Senn, W. Somato-dendritic synaptic plasticity and error-backpropagation in active dendrites. PLoS Comput. Biol. 12 , e1004638 (2016).
Poirazi, P. & Papoutsi, A. Illuminating dendritic function with computational models. Nat. Rev. Neurosci. 21 , 303–321 (2020).
Beniaguev, D., Segev, I. & London, M. Single cortical neurons as deep artificial neural networks. Neuron 109 , 2727–2739.e3 (2021).
Cossell, L. et al. Functional organization of excitatory synaptic strength in primary visual cortex. Nature 518 , 399–403 (2015).
Seeman, S. C. et al. Sparse recurrent excitatory connectivity in the microcircuit of the adult mouse and human cortex. eLife 7 , e37349 (2018).
Garner, A. R. & Keller, G. B. A cortical circuit for audio-visual predictions. Nat. Neurosci. 25 , 98–105 (2022).
Ghazanfar, A. A. & Schroeder, C. E. Is neocortex essentially multisensory? Trends Cogn. Sci. 10 , 278–285 (2006).
Fetsch, C. R., DeAngelis, G. C. & Angelaki, D. E. Bridging the gap between theories of sensory cue integration and the physiology of multisensory neurons. Nat. Rev. Neurosci. 14 , 429–442 (2013).
Graybiel, A. M. The basal ganglia. Curr. Biol. 10 , R509–R511 (2000).
Alexander, G. E., DeLong, M. R. & Strick, P. L. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annu. Rev. Neurosci. 9 , 357–381 (1986).
Parent, A. et al. Organization of the basal ganglia: the importance of axonal collateralization. Trends Neurosci. 23 , S20–S27 (2000).
Takakusaki, K., Saitoh, K., Harada, H. & Kashiwayanagi, M. Role of basal ganglia–brainstem pathways in the control of motor behaviors. Neurosci. Res. 50 , 137–151 (2004).
Graybiel, A. M., Aosaki, T., Flaherty, A. W. & Kimura, M. The basal ganglia and adaptive motor control. Science 265 , 1826–1831 (1994).
Roseberry, T. K. et al. Cell-type-specific control of brainstem locomotor circuits by basal ganglia. Cell 164 , 526–537 (2016).
Parent, M., Levesque, M. & Parent, A. Two types of projection neurons in the internal pallidum of primates: single-axon tracing and three-dimensional reconstruction. J. Comp. Neurol. 439 , 162–175 (2001).
Parent, M. & Parent, A. The pallidofugal motor fiber system in primates. Parkinsonism Relat. Disord. 10 , 203–211 (2004).
Pennartz, C. M., Groenewegen, H. J. & Lopes da Silva, F. H. The nucleus accumbens as a complex of functionally distinct neuronal ensembles: an integration of behavioural, electrophysiological and anatomical data. Prog. Neurobiol. 42 , 719–761 (1994).
Di Chiara, G., Porceddu, M. L., Morelli, M., Mulas, M. L. & Gessa, G. L. Evidence for a GABAergic projection from the substantia nigra to the ventromedial thalamus and to the superior colliculus of the rat. Brain Res. 176 , 273–284 (1979).
Williams, L. E. & Holtmaat, A. Higher-order thalamocortical inputs gate synaptic long-term potentiation via disinhibition. Neuron 101 , 91–102.e4 (2019).
Gambino, F. et al. Sensory-evoked LTP driven by dendritic plateau potentials in vivo. Nature 515 , 116–119 (2014).
Anastasiades, P. G., Collins, D. P. & Carter, A. G. Mediodorsal and ventromedial thalamus engage distinct L1 circuits in the prefrontal cortex. Neuron 109 , 314–330.e4 (2021).
Schmitt, L. I. et al. Thalamic amplification of cortical connectivity sustains attentional control. Nature 545 , 219–223 (2017).
Inagaki, H. K. et al. A midbrain–thalamus–cortex circuit reorganizes cortical dynamics to initiate movement. Cell 185 , 1065–1081.e23 (2022).
Wang, M. B. & Halassa, M. M. Thalamocortical contribution to flexible learning in neural systems. Netw. Neurosci. 6 , 980–997 (2022).
La Terra, D. et al. The role of higher-order thalamus during learning and correct performance in goal-directed behavior. eLife 11 , e77177 (2022).
Ruis, L., Andreas, J., Baroni, M., Bouchacourt, D. & Lake, B. M. A benchmark for systematic generalization in grounded language understanding. In Proc. 34th Int. Conf. Neural Information Processing Systems (eds. Larochelle, H. et al.) 19861–19872 (Curran, 2020).
Lake, B. M. & Baroni, M. Generalization without systematicity: on the compositional skills of sequence-to-sequence recurrent networks. In Int. Conf. Machine Learning (eds. Dy, J. & Krause, A.) 2879–2888 (2018).
Pfeiffer, J., Ruder, S., Vulić, I. & Ponti, E. M. Modular deep learning. Preprint at arXiv https://doi.org/10.48550/arXiv.2302.11529 (2023).
Goyal, A. et al. Recurrent independent mechanisms. Preprint at arXiv https://doi.org/10.48550/arXiv.1909.10893 (2020).
Albright, T. D., Jessell, T. M., Kandel, E. R. & Posner, M. I. Neural science: a century of progress and the mysteries that remain. Neuron 25 , S1–S55 (2000).
Wallis, J. D., Anderson, K. C. & Miller, E. K. Single neurons in prefrontal cortex encode abstract rules. Nature 411 , 953–956 (2001).
Verschure, P. F., Pennartz, C. M. & Pezzulo, G. The why, what, where, when and how of goal-directed choice: neuronal and computational principles. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369 , 20130483 (2014).
Dias, R., Robbins, T. W. & Roberts, A. C. Dissociation in prefrontal cortex of affective and attentional shifts. Nature 380 , 69–72 (1996).
Wilson, R. C., Takahashi, Y. K., Schoenbaum, G. & Niv, Y. Orbitofrontal cortex as a cognitive map of task space. Neuron 81 , 267–279 (2014).
Fan, J., McCandliss, B. D., Fossella, J., Flombaum, J. I. & Posner, M. I. The activation of attentional networks. Neuroimage 26 , 471–479 (2005).
Womelsdorf, T. & Everling, S. Long-range attention networks: circuit motifs underlying endogenously controlled stimulus selection. Trends Neurosci. 38 , 682–700 (2015).
Cohen, M. R. & Maunsell, J. H. Attention improves performance primarily by reducing interneuronal correlations. Nat. Neurosci. 12 , 1594–1600 (2009).
Reynolds, J. H. & Desimone, R. Interacting roles of attention and visual salience in V4. Neuron 37 , 853–863 (2003).
Poort, J. et al. The role of attention in figure-ground segregation in areas V1 and V4 of the visual cortex. Neuron 75 , 143–156 (2012).
Reep, R. L. & Corwin, J. V. Posterior parietal cortex as part of a neural network for directed attention in rats. Neurobiol. Learn. Mem. 91 , 104–113 (2009).
Saalmann, Y. B., Pinsk, M. A., Wang, L., Li, X. & Kastner, S. The pulvinar regulates information transmission between cortical areas based on attention demands. Science 337 , 753–756 (2012).
Rikhye, R. V., Gilra, A. & Halassa, M. M. Thalamic regulation of switching between cortical representations enables cognitive flexibility. Nat. Neurosci. 21 , 1753–1763 (2018).
Van der Werf, Y. D., Witter, M. P. & Groenewegen, H. J. The intralaminar and midline nuclei of the thalamus. Anatomical and functional evidence for participation in processes of arousal and awareness. Brain Res. Brain Res. Rev. 39 , 107–140 (2002).
Groenewegen, H. J. & Berendse, H. W. The specificity of the ‘nonspecific’ midline and intralaminar thalamic nuclei. Trends Neurosci. 17 , 52–57 (1994).
Breton-Provencher, V., Drummond, G. T., Feng, J., Li, Y. & Sur, M. Spatiotemporal dynamics of noradrenaline during learned behaviour. Nature 606 , 732–738 (2022).
Ren, J. et al. Anatomically defined and functionally distinct dorsal raphe serotonin sub-systems. Cell 175 , 472–487.e20 (2018).
Lohani, S. et al. Spatiotemporally heterogeneous coordination of cholinergic and neocortical activity. Nat. Neurosci. 25 , 1706–1713 (2022).
Morris, L. S. et al. Fronto-striatal organization: defining functional and microstructural substrates of behavioural flexibility. Cortex 74 , 118–133 (2016).
Apicella, P., Legallet, E., Nieoullon, A. & Trouche, E. Neglect of contralateral visual stimuli in monkeys with unilateral striatal dopamine depletion. Behav. Brain Res. 46 , 187–195 (1991).
Download references
Acknowledgements
M.S. discloses support for the research of this work from the Brain Science Foundation and the Sumitomo Foundation (2200084). C.M.A.P. discloses support for the research of this work from the European Union’s Horizon 2020 Framework Program for Research and Innovation (Human Brain Project SGA3, 945539). J.A. discloses support for the research of this work from the European Social Fund through the ‘ICT programme’ measure and the Estonian Research Council grant (PSG728).
Author information
Authors and affiliations.
Department of Cognitive and Systems Neuroscience, Swammerdam Institute for Life Sciences, University of Amsterdam, Amsterdam, The Netherlands
Mototaka Suzuki & Cyriel M. A. Pennartz
Institute of Computer Science, University of Tartu, Tartu, Estonia
You can also search for this author in PubMed Google Scholar
Contributions
The authors contributed equally to all aspects of the article.
Corresponding authors
Correspondence to Mototaka Suzuki or Jaan Aru .
Ethics declarations
Competing interests.
The authors declare no competing interests.
Peer review
Peer review information.
Nature Reviews Neuroscience thanks Andreas Burkhalter and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
A method of statistical analysis that is grounded in Bayes’ theorem, which describes how the probability of a hypothesis (posterior probability) is updated as new data (evidence) become available, given prior knowledge about the hypothesis (prior probability).
Neural circuits that connect different regions of the cerebral cortex to one another, allowing communication and integration of information across various cortical areas. These loops can be either short range, connecting adjacent or nearby cortical regions, or long range, linking distant regions of the cortex.
A hierarchical structure consisting of many layers (roughly analogous to cortical areas) through which information from the external world is processed step by step.
Structured configurations of hierarchical, interconnected layers of artificial neurons, or nodes, in a neural network. Common types of deep learning architecture include feedforward convolutional neural networks and recurrent neural networks.
The process of drawing conclusions from data wherein parameters are organized into different levels or layers. In hierarchical Bayesian inference, Bayesian statistics are employed within a layered framework, integrating prior knowledge at multiple levels to refine posterior distributions.
Thalamic nuclei can be categorized anatomically into first-order and higher-order nuclei. First-order nuclei receive driving afferents from ascending pathways, whereas the higher-order nuclei receive driving afferents from cortical layer 5 pyramidal (L5p) neurons. Notable examples of higher-order thalamic nuclei include the pulvinar and the medial dorsal nucleus.
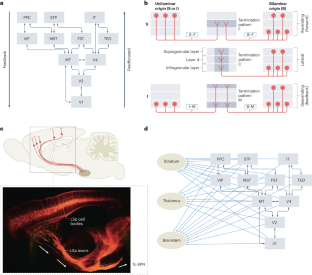
Connections made between two cortical areas that are not distinguished hierarchically (for instance, primary auditory and visual cortex). This connectivity pattern is illustrated in Fig. 1b .
Connections in which the output of a neuron at a given layer is fed back as an input to either the same layer or a previous layer. This creates a loop in the network, allowing information, for instance, to persist and be reused across sequential steps.
A class of neural networks in which connections between nodes form directed cycles, enabling the retention of information from previous inputs. This sequential memory feature makes recurrent neural networks suitable for tasks involving time-series or sequential data.
A machine learning method in which an agent makes decisions and receives reinforcing feedback to train the network to improve its output (for example, reward for desired behaviours, punishment for behaviour resulting in undesirable output).
Architectures that do not consist of a deep hierarchy. Shallow architectures instead have a minimum number of layers.
Computations carried out by a shallow architecture, namely in a few steps instead of tens or hundreds of layers of processing.
Bidirectional pathways between the thalamus and the cerebral cortex. Thalamo-cortical loops play a vital role in the regulation of consciousness, attention and sensory processing, and have been implicated in several neurological and psychiatric disorders.
Connections made between two brain regions via the thalamus.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
Reprints and permissions
About this article
Cite this article.
Suzuki, M., Pennartz, C.M.A. & Aru, J. How deep is the brain? The shallow brain hypothesis. Nat. Rev. Neurosci. 24 , 778–791 (2023). https://doi.org/10.1038/s41583-023-00756-z
Download citation
Accepted : 25 September 2023
Published : 27 October 2023
Issue Date : December 2023
DOI : https://doi.org/10.1038/s41583-023-00756-z
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
Quick links
- Explore articles by subject
- Guide to authors
- Editorial policies
Sign up for the Nature Briefing newsletter — what matters in science, free to your inbox daily.

- Search Menu
- Browse content in Arts and Humanities
- Browse content in Archaeology
- Anglo-Saxon and Medieval Archaeology
- Archaeological Methodology and Techniques
- Archaeology by Region
- Archaeology of Religion
- Archaeology of Trade and Exchange
- Biblical Archaeology
- Contemporary and Public Archaeology
- Environmental Archaeology
- Historical Archaeology
- History and Theory of Archaeology
- Industrial Archaeology
- Landscape Archaeology
- Mortuary Archaeology
- Prehistoric Archaeology
- Underwater Archaeology
- Urban Archaeology
- Zooarchaeology
- Browse content in Architecture
- Architectural Structure and Design
- History of Architecture
- Residential and Domestic Buildings
- Theory of Architecture
- Browse content in Art
- Art Subjects and Themes
- History of Art
- Industrial and Commercial Art
- Theory of Art
- Biographical Studies
- Byzantine Studies
- Browse content in Classical Studies
- Classical History
- Classical Philosophy
- Classical Mythology
- Classical Literature
- Classical Reception
- Classical Art and Architecture
- Classical Oratory and Rhetoric
- Greek and Roman Epigraphy
- Greek and Roman Law
- Greek and Roman Papyrology
- Greek and Roman Archaeology
- Late Antiquity
- Religion in the Ancient World
- Digital Humanities
- Browse content in History
- Colonialism and Imperialism
- Diplomatic History
- Environmental History
- Genealogy, Heraldry, Names, and Honours
- Genocide and Ethnic Cleansing
- Historical Geography
- History by Period
- History of Emotions
- History of Agriculture
- History of Education
- History of Gender and Sexuality
- Industrial History
- Intellectual History
- International History
- Labour History
- Legal and Constitutional History
- Local and Family History
- Maritime History
- Military History
- National Liberation and Post-Colonialism
- Oral History
- Political History
- Public History
- Regional and National History
- Revolutions and Rebellions
- Slavery and Abolition of Slavery
- Social and Cultural History
- Theory, Methods, and Historiography
- Urban History
- World History
- Browse content in Language Teaching and Learning
- Language Learning (Specific Skills)
- Language Teaching Theory and Methods
- Browse content in Linguistics
- Applied Linguistics
- Cognitive Linguistics
- Computational Linguistics
- Forensic Linguistics
- Grammar, Syntax and Morphology
- Historical and Diachronic Linguistics
- History of English
- Language Acquisition
- Language Evolution
- Language Reference
- Language Variation
- Language Families
- Lexicography
- Linguistic Anthropology
- Linguistic Theories
- Linguistic Typology
- Phonetics and Phonology
- Psycholinguistics
- Sociolinguistics
- Translation and Interpretation
- Writing Systems
- Browse content in Literature
- Bibliography
- Children's Literature Studies
- Literary Studies (Asian)
- Literary Studies (European)
- Literary Studies (Eco-criticism)
- Literary Studies (Romanticism)
- Literary Studies (American)
- Literary Studies (Modernism)
- Literary Studies - World
- Literary Studies (1500 to 1800)
- Literary Studies (19th Century)
- Literary Studies (20th Century onwards)
- Literary Studies (African American Literature)
- Literary Studies (British and Irish)
- Literary Studies (Early and Medieval)
- Literary Studies (Fiction, Novelists, and Prose Writers)
- Literary Studies (Gender Studies)
- Literary Studies (Graphic Novels)
- Literary Studies (History of the Book)
- Literary Studies (Plays and Playwrights)
- Literary Studies (Poetry and Poets)
- Literary Studies (Postcolonial Literature)
- Literary Studies (Queer Studies)
- Literary Studies (Science Fiction)
- Literary Studies (Travel Literature)
- Literary Studies (War Literature)
- Literary Studies (Women's Writing)
- Literary Theory and Cultural Studies
- Mythology and Folklore
- Shakespeare Studies and Criticism
- Browse content in Media Studies
- Browse content in Music
- Applied Music
- Dance and Music
- Ethics in Music
- Ethnomusicology
- Gender and Sexuality in Music
- Medicine and Music
- Music Cultures
- Music and Religion
- Music and Media
- Music and Culture
- Music Education and Pedagogy
- Music Theory and Analysis
- Musical Scores, Lyrics, and Libretti
- Musical Structures, Styles, and Techniques
- Musicology and Music History
- Performance Practice and Studies
- Race and Ethnicity in Music
- Sound Studies
- Browse content in Performing Arts
- Browse content in Philosophy
- Aesthetics and Philosophy of Art
- Epistemology
- Feminist Philosophy
- History of Western Philosophy
- Metaphysics
- Moral Philosophy
- Non-Western Philosophy
- Philosophy of Science
- Philosophy of Language
- Philosophy of Mind
- Philosophy of Perception
- Philosophy of Action
- Philosophy of Law
- Philosophy of Religion
- Philosophy of Mathematics and Logic
- Practical Ethics
- Social and Political Philosophy
- Browse content in Religion
- Biblical Studies
- Christianity
- East Asian Religions
- History of Religion
- Judaism and Jewish Studies
- Qumran Studies
- Religion and Education
- Religion and Health
- Religion and Politics
- Religion and Science
- Religion and Law
- Religion and Art, Literature, and Music
- Religious Studies
- Browse content in Society and Culture
- Cookery, Food, and Drink
- Cultural Studies
- Customs and Traditions
- Ethical Issues and Debates
- Hobbies, Games, Arts and Crafts
- Lifestyle, Home, and Garden
- Natural world, Country Life, and Pets
- Popular Beliefs and Controversial Knowledge
- Sports and Outdoor Recreation
- Technology and Society
- Travel and Holiday
- Visual Culture
- Browse content in Law
- Arbitration
- Browse content in Company and Commercial Law
- Commercial Law
- Company Law
- Browse content in Comparative Law
- Systems of Law
- Competition Law
- Browse content in Constitutional and Administrative Law
- Government Powers
- Judicial Review
- Local Government Law
- Military and Defence Law
- Parliamentary and Legislative Practice
- Construction Law
- Contract Law
- Browse content in Criminal Law
- Criminal Procedure
- Criminal Evidence Law
- Sentencing and Punishment
- Employment and Labour Law
- Environment and Energy Law
- Browse content in Financial Law
- Banking Law
- Insolvency Law
- History of Law
- Human Rights and Immigration
- Intellectual Property Law
- Browse content in International Law
- Private International Law and Conflict of Laws
- Public International Law
- IT and Communications Law
- Jurisprudence and Philosophy of Law
- Law and Politics
- Law and Society
- Browse content in Legal System and Practice
- Courts and Procedure
- Legal Skills and Practice
- Primary Sources of Law
- Regulation of Legal Profession
- Medical and Healthcare Law
- Browse content in Policing
- Criminal Investigation and Detection
- Police and Security Services
- Police Procedure and Law
- Police Regional Planning
- Browse content in Property Law
- Personal Property Law
- Study and Revision
- Terrorism and National Security Law
- Browse content in Trusts Law
- Wills and Probate or Succession
- Browse content in Medicine and Health
- Browse content in Allied Health Professions
- Arts Therapies
- Clinical Science
- Dietetics and Nutrition
- Occupational Therapy
- Operating Department Practice
- Physiotherapy
- Radiography
- Speech and Language Therapy
- Browse content in Anaesthetics
- General Anaesthesia
- Neuroanaesthesia
- Browse content in Clinical Medicine
- Acute Medicine
- Cardiovascular Medicine
- Clinical Genetics
- Clinical Pharmacology and Therapeutics
- Dermatology
- Endocrinology and Diabetes
- Gastroenterology
- Genito-urinary Medicine
- Geriatric Medicine
- Infectious Diseases
- Medical Toxicology
- Medical Oncology
- Pain Medicine
- Palliative Medicine
- Rehabilitation Medicine
- Respiratory Medicine and Pulmonology
- Rheumatology
- Sleep Medicine
- Sports and Exercise Medicine
- Clinical Neuroscience
- Community Medical Services
- Critical Care
- Emergency Medicine
- Forensic Medicine
- Haematology
- History of Medicine
- Browse content in Medical Dentistry
- Oral and Maxillofacial Surgery
- Paediatric Dentistry
- Restorative Dentistry and Orthodontics
- Surgical Dentistry
- Browse content in Medical Skills
- Clinical Skills
- Communication Skills
- Nursing Skills
- Surgical Skills
- Medical Ethics
- Medical Statistics and Methodology
- Browse content in Neurology
- Clinical Neurophysiology
- Neuropathology
- Nursing Studies
- Browse content in Obstetrics and Gynaecology
- Gynaecology
- Occupational Medicine
- Ophthalmology
- Otolaryngology (ENT)
- Browse content in Paediatrics
- Neonatology
- Browse content in Pathology
- Chemical Pathology
- Clinical Cytogenetics and Molecular Genetics
- Histopathology
- Medical Microbiology and Virology
- Patient Education and Information
- Browse content in Pharmacology
- Psychopharmacology
- Browse content in Popular Health
- Caring for Others
- Complementary and Alternative Medicine
- Self-help and Personal Development
- Browse content in Preclinical Medicine
- Cell Biology
- Molecular Biology and Genetics
- Reproduction, Growth and Development
- Primary Care
- Professional Development in Medicine
- Browse content in Psychiatry
- Addiction Medicine
- Child and Adolescent Psychiatry
- Forensic Psychiatry
- Learning Disabilities
- Old Age Psychiatry
- Psychotherapy
- Browse content in Public Health and Epidemiology
- Epidemiology
- Public Health
- Browse content in Radiology
- Clinical Radiology
- Interventional Radiology
- Nuclear Medicine
- Radiation Oncology
- Reproductive Medicine
- Browse content in Surgery
- Cardiothoracic Surgery
- Gastro-intestinal and Colorectal Surgery
- General Surgery
- Neurosurgery
- Paediatric Surgery
- Peri-operative Care
- Plastic and Reconstructive Surgery
- Surgical Oncology
- Transplant Surgery
- Trauma and Orthopaedic Surgery
- Vascular Surgery
- Browse content in Science and Mathematics
- Browse content in Biological Sciences
- Aquatic Biology
- Biochemistry
- Bioinformatics and Computational Biology
- Developmental Biology
- Ecology and Conservation
- Evolutionary Biology
- Genetics and Genomics
- Microbiology
- Molecular and Cell Biology
- Natural History
- Plant Sciences and Forestry
- Research Methods in Life Sciences
- Structural Biology
- Systems Biology
- Zoology and Animal Sciences
- Browse content in Chemistry
- Analytical Chemistry
- Computational Chemistry
- Crystallography
- Environmental Chemistry
- Industrial Chemistry
- Inorganic Chemistry
- Materials Chemistry
- Medicinal Chemistry
- Mineralogy and Gems
- Organic Chemistry
- Physical Chemistry
- Polymer Chemistry
- Study and Communication Skills in Chemistry
- Theoretical Chemistry
- Browse content in Computer Science
- Artificial Intelligence
- Computer Architecture and Logic Design
- Game Studies
- Human-Computer Interaction
- Mathematical Theory of Computation
- Programming Languages
- Software Engineering
- Systems Analysis and Design
- Virtual Reality
- Browse content in Computing
- Business Applications
- Computer Security
- Computer Games
- Computer Networking and Communications
- Digital Lifestyle
- Graphical and Digital Media Applications
- Operating Systems
- Browse content in Earth Sciences and Geography
- Atmospheric Sciences
- Environmental Geography
- Geology and the Lithosphere
- Maps and Map-making
- Meteorology and Climatology
- Oceanography and Hydrology
- Palaeontology
- Physical Geography and Topography
- Regional Geography
- Soil Science
- Urban Geography
- Browse content in Engineering and Technology
- Agriculture and Farming
- Biological Engineering
- Civil Engineering, Surveying, and Building
- Electronics and Communications Engineering
- Energy Technology
- Engineering (General)
- Environmental Science, Engineering, and Technology
- History of Engineering and Technology
- Mechanical Engineering and Materials
- Technology of Industrial Chemistry
- Transport Technology and Trades
- Browse content in Environmental Science
- Applied Ecology (Environmental Science)
- Conservation of the Environment (Environmental Science)
- Environmental Sustainability
- Environmentalist Thought and Ideology (Environmental Science)
- Management of Land and Natural Resources (Environmental Science)
- Natural Disasters (Environmental Science)
- Nuclear Issues (Environmental Science)
- Pollution and Threats to the Environment (Environmental Science)
- Social Impact of Environmental Issues (Environmental Science)
- History of Science and Technology
- Browse content in Materials Science
- Ceramics and Glasses
- Composite Materials
- Metals, Alloying, and Corrosion
- Nanotechnology
- Browse content in Mathematics
- Applied Mathematics
- Biomathematics and Statistics
- History of Mathematics
- Mathematical Education
- Mathematical Finance
- Mathematical Analysis
- Numerical and Computational Mathematics
- Probability and Statistics
- Pure Mathematics
- Browse content in Neuroscience
- Cognition and Behavioural Neuroscience
- Development of the Nervous System
- Disorders of the Nervous System
- History of Neuroscience
- Invertebrate Neurobiology
- Molecular and Cellular Systems
- Neuroendocrinology and Autonomic Nervous System
- Neuroscientific Techniques
- Sensory and Motor Systems
- Browse content in Physics
- Astronomy and Astrophysics
- Atomic, Molecular, and Optical Physics
- Biological and Medical Physics
- Classical Mechanics
- Computational Physics
- Condensed Matter Physics
- Electromagnetism, Optics, and Acoustics
- History of Physics
- Mathematical and Statistical Physics
- Measurement Science
- Nuclear Physics
- Particles and Fields
- Plasma Physics
- Quantum Physics
- Relativity and Gravitation
- Semiconductor and Mesoscopic Physics
- Browse content in Psychology
- Affective Sciences
- Clinical Psychology
- Cognitive Psychology
- Cognitive Neuroscience
- Criminal and Forensic Psychology
- Developmental Psychology
- Educational Psychology
- Evolutionary Psychology
- Health Psychology
- History and Systems in Psychology
- Music Psychology
- Neuropsychology
- Organizational Psychology
- Psychological Assessment and Testing
- Psychology of Human-Technology Interaction
- Psychology Professional Development and Training
- Research Methods in Psychology
- Social Psychology
- Browse content in Social Sciences
- Browse content in Anthropology
- Anthropology of Religion
- Human Evolution
- Medical Anthropology
- Physical Anthropology
- Regional Anthropology
- Social and Cultural Anthropology
- Theory and Practice of Anthropology
- Browse content in Business and Management
- Business Strategy
- Business Ethics
- Business History
- Business and Government
- Business and Technology
- Business and the Environment
- Comparative Management
- Corporate Governance
- Corporate Social Responsibility
- Entrepreneurship
- Health Management
- Human Resource Management
- Industrial and Employment Relations
- Industry Studies
- Information and Communication Technologies
- International Business
- Knowledge Management
- Management and Management Techniques
- Operations Management
- Organizational Theory and Behaviour
- Pensions and Pension Management
- Public and Nonprofit Management
- Strategic Management
- Supply Chain Management
- Browse content in Criminology and Criminal Justice
- Criminal Justice
- Criminology
- Forms of Crime
- International and Comparative Criminology
- Youth Violence and Juvenile Justice
- Development Studies
- Browse content in Economics
- Agricultural, Environmental, and Natural Resource Economics
- Asian Economics
- Behavioural Finance
- Behavioural Economics and Neuroeconomics
- Econometrics and Mathematical Economics
- Economic Systems
- Economic History
- Economic Methodology
- Economic Development and Growth
- Financial Markets
- Financial Institutions and Services
- General Economics and Teaching
- Health, Education, and Welfare
- History of Economic Thought
- International Economics
- Labour and Demographic Economics
- Law and Economics
- Macroeconomics and Monetary Economics
- Microeconomics
- Public Economics
- Urban, Rural, and Regional Economics
- Welfare Economics
- Browse content in Education
- Adult Education and Continuous Learning
- Care and Counselling of Students
- Early Childhood and Elementary Education
- Educational Equipment and Technology
- Educational Strategies and Policy
- Higher and Further Education
- Organization and Management of Education
- Philosophy and Theory of Education
- Schools Studies
- Secondary Education
- Teaching of a Specific Subject
- Teaching of Specific Groups and Special Educational Needs
- Teaching Skills and Techniques
- Browse content in Environment
- Applied Ecology (Social Science)
- Climate Change
- Conservation of the Environment (Social Science)
- Environmentalist Thought and Ideology (Social Science)
- Natural Disasters (Environment)
- Social Impact of Environmental Issues (Social Science)
- Browse content in Human Geography
- Cultural Geography
- Economic Geography
- Political Geography
- Browse content in Interdisciplinary Studies
- Communication Studies
- Museums, Libraries, and Information Sciences
- Browse content in Politics
- African Politics
- Asian Politics
- Chinese Politics
- Comparative Politics
- Conflict Politics
- Elections and Electoral Studies
- Environmental Politics
- European Union
- Foreign Policy
- Gender and Politics
- Human Rights and Politics
- Indian Politics
- International Relations
- International Organization (Politics)
- International Political Economy
- Irish Politics
- Latin American Politics
- Middle Eastern Politics
- Political Methodology
- Political Communication
- Political Philosophy
- Political Sociology
- Political Behaviour
- Political Economy
- Political Institutions
- Political Theory
- Politics and Law
- Public Administration
- Public Policy
- Quantitative Political Methodology
- Regional Political Studies
- Russian Politics
- Security Studies
- State and Local Government
- UK Politics
- US Politics
- Browse content in Regional and Area Studies
- African Studies
- Asian Studies
- East Asian Studies
- Japanese Studies
- Latin American Studies
- Middle Eastern Studies
- Native American Studies
- Scottish Studies
- Browse content in Research and Information
- Research Methods
- Browse content in Social Work
- Addictions and Substance Misuse
- Adoption and Fostering
- Care of the Elderly
- Child and Adolescent Social Work
- Couple and Family Social Work
- Developmental and Physical Disabilities Social Work
- Direct Practice and Clinical Social Work
- Emergency Services
- Human Behaviour and the Social Environment
- International and Global Issues in Social Work
- Mental and Behavioural Health
- Social Justice and Human Rights
- Social Policy and Advocacy
- Social Work and Crime and Justice
- Social Work Macro Practice
- Social Work Practice Settings
- Social Work Research and Evidence-based Practice
- Welfare and Benefit Systems
- Browse content in Sociology
- Childhood Studies
- Community Development
- Comparative and Historical Sociology
- Economic Sociology
- Gender and Sexuality
- Gerontology and Ageing
- Health, Illness, and Medicine
- Marriage and the Family
- Migration Studies
- Occupations, Professions, and Work
- Organizations
- Population and Demography
- Race and Ethnicity
- Social Theory
- Social Movements and Social Change
- Social Research and Statistics
- Social Stratification, Inequality, and Mobility
- Sociology of Religion
- Sociology of Education
- Sport and Leisure
- Urban and Rural Studies
- Browse content in Warfare and Defence
- Defence Strategy, Planning, and Research
- Land Forces and Warfare
- Military Administration
- Military Life and Institutions
- Naval Forces and Warfare
- Other Warfare and Defence Issues
- Peace Studies and Conflict Resolution
- Weapons and Equipment

- < Previous chapter
- Next chapter >

3 The Social Brain Hypothesis: An Evolutionary Perspective on the Neurobiology of Social Behaviour
- Published: February 2014
- Cite Icon Cite
- Permissions Icon Permissions
This chapter places human cognition into an evolutionary context by describing brain and cognitive evolution in humans' closest relatives, the primates (prosimians, monkeys, and apes), and how human cognition differs from that of other animals. In particular, it examines ‘social cognition’, the kinds of cognitive processes that are believed to underlie social behaviour and social relationships. After reviewing what is known about social cognition in the brain and what insights neuroimaging has provided about how humans execute socio-cognitive tasks, the chapter discusses the contribution that has been made by the rapid advances in neuroimaging technology during the last decade or so. Finally, it explores the main assumptions of the social brain hypothesis, which attributes the large brains of primates to the cognitive demands imposed by social complexity.
Signed in as
Institutional accounts.
- Google Scholar Indexing
- GoogleCrawler [DO NOT DELETE]
Personal account
- Sign in with email/username & password
- Get email alerts
- Save searches
- Purchase content
- Activate your purchase/trial code
Institutional access
- Sign in with a library card Sign in with username/password Recommend to your librarian
- Institutional account management
- Get help with access
Access to content on Oxford Academic is often provided through institutional subscriptions and purchases. If you are a member of an institution with an active account, you may be able to access content in one of the following ways:
IP based access
Typically, access is provided across an institutional network to a range of IP addresses. This authentication occurs automatically, and it is not possible to sign out of an IP authenticated account.
Sign in through your institution
Choose this option to get remote access when outside your institution. Shibboleth/Open Athens technology is used to provide single sign-on between your institution’s website and Oxford Academic.
- Click Sign in through your institution.
- Select your institution from the list provided, which will take you to your institution's website to sign in.
- When on the institution site, please use the credentials provided by your institution. Do not use an Oxford Academic personal account.
- Following successful sign in, you will be returned to Oxford Academic.
If your institution is not listed or you cannot sign in to your institution’s website, please contact your librarian or administrator.
Sign in with a library card
Enter your library card number to sign in. If you cannot sign in, please contact your librarian.
Society Members
Society member access to a journal is achieved in one of the following ways:
Sign in through society site
Many societies offer single sign-on between the society website and Oxford Academic. If you see ‘Sign in through society site’ in the sign in pane within a journal:
- Click Sign in through society site.
- When on the society site, please use the credentials provided by that society. Do not use an Oxford Academic personal account.
If you do not have a society account or have forgotten your username or password, please contact your society.
Sign in using a personal account
Some societies use Oxford Academic personal accounts to provide access to their members. See below.
A personal account can be used to get email alerts, save searches, purchase content, and activate subscriptions.
Some societies use Oxford Academic personal accounts to provide access to their members.
Viewing your signed in accounts
Click the account icon in the top right to:
- View your signed in personal account and access account management features.
- View the institutional accounts that are providing access.
Signed in but can't access content
Oxford Academic is home to a wide variety of products. The institutional subscription may not cover the content that you are trying to access. If you believe you should have access to that content, please contact your librarian.
For librarians and administrators, your personal account also provides access to institutional account management. Here you will find options to view and activate subscriptions, manage institutional settings and access options, access usage statistics, and more.
Our books are available by subscription or purchase to libraries and institutions.
- About Oxford Academic
- Publish journals with us
- University press partners
- What we publish
- New features
- Open access
- Rights and permissions
- Accessibility
- Advertising
- Media enquiries
- Oxford University Press
- Oxford Languages
- University of Oxford
Oxford University Press is a department of the University of Oxford. It furthers the University's objective of excellence in research, scholarship, and education by publishing worldwide
- Copyright © 2024 Oxford University Press
- Cookie settings
- Cookie policy
- Privacy policy
- Legal notice
This Feature Is Available To Subscribers Only
Sign In or Create an Account
This PDF is available to Subscribers Only
For full access to this pdf, sign in to an existing account, or purchase an annual subscription.

Authenticity
Let’s not try to be “authentic”, instead of authenticity, think about narrating your own life journey..
Posted March 29, 2024 | Reviewed by Tyler Woods
- Our core selves change throughout our lives.
- The true “self” is a moving target.
- We can narrative the stories of our lives to understand ourselves.

One of the big recent cultural shifts in America has been people claiming that they want to strive toward greater authenticity in their lives. Perhaps this was brought on by a feeling that people are increasingly “fake” in their online personas, especially on social media and dating app profiles, or in routine interactions with acquaintances or colleagues that might seem forced and mundane. Whatever the reason, people seem to be strongly craving a connection with their true selves and to bring more authenticity into their lives.
There’s just one problem. There is no true self, at least not in any sense of the self that we can understand through science. We should seriously question the idea of authenticity as a meaningful construct in our lives.
A Psychological Quirk
We might naively assume that everyone has a true and authentic core “self,” almost like believing in a secular version of a soul. But this assumption is based on a psychological quirk. Humans are essentialist thinkers, which means we wrongly assume that all beings have a stable underlying essence, or a je ne sais quoi . But when it comes to living, growing life forms, this is an illusion. Lots of people say that in order to have a happy, meaningful life, we just need to get in touch with that core, essentialist part of ourselves and behave in ways that are consistent with this. I suggest this is a false and unhealthy way of thinking. This concept of authenticity isn’t useful because it’s based on a flawed assumption of how human psychology works.
Previously , I suggested that people actually change a lot throughout their lives. But the pace of change diminishes throughout adulthood, so it may not feel subjectively like we’re changing very much from year to year when, in fact, we do. This also helps explain why it may not be possible to have one true authentic self, because the self is almost always changing. Striving for authenticity may be like trying to hit a moving target. If you’re always in motion, or if your north star keeps shifting, then getting to an authentic state would be a fool’s errand. Developmental psychology is, in a sense, the study of change. The more we change, the more it should be apparent to us that there isn’t a fixed, static thing that we should use to define ourselves.
One might think that this would be a liberating idea. With so much in flux, life has seemingly endless possibilities! Being truly authentic might be like going on a never-ending scavenger hunt. You might spend your life looking for elements of yourself (which can be really fun!) even if you never find all of them. The idea of one "true" self could be too restricting.
We can also imagine the opposite, that the idea of moving through life without a well-defined, core sense of self could feel destabilizing or even anxiety -provoking. This is why people often crave certainty and stability. This is sometimes referred to as a "need for cognitive closure." People want to know how things in the world really do work, now just how they may or may not work. Just like everything else, we want the same level of insight and understanding into ourselves.
Narrating Your Own Story
Another way to reconcile these ideas is through a theory of personality development championed by researcher Dan McAdams. His research suggests that our personalities change based on adaptations. You might begin your life as a more introverted person, but then meet friends in young adulthood who pull you out of your shell. Thus, you become a more extroverted person through those social relationships. So, which was the “authentic” version of yourself: the initial introvert or the eventual extrovert?
McAdams suggests that both are real, true parts of yourself, and the way we make sense of this is to tell a story about our lives that weaves all of these threads together. By narrating the story of our own journeys, we behave as what McAdams calls the "autobiographical author." We tell the story of our lives, including how we evolve and adapt to different life circumstances (sometimes permanently). This is, in my view, a better way that people can attempt to be “authentic.” I suggest that developing one's own narrative-based identity is a much healthier way for people to get in tune with their own selves.
McAdams, D. P. (2013). The psychological self as actor, agent, and author. Perspectives on Psychological Science, 8 (3), 272-295.

Dylan Selterman, Ph.D., is an Associate Teaching Professor at Johns Hopkins University in the Department of Psychological and Brain Sciences. He teaches courses and conducts research on personality traits, happiness, relationships, morality/ethics, game theory, political psychology, and more.
- Find a Therapist
- Find a Treatment Center
- Find a Support Group
- International
- New Zealand
- South Africa
- Switzerland
- Asperger's
- Bipolar Disorder
- Chronic Pain
- Eating Disorders
- Passive Aggression
- Personality
- Goal Setting
- Positive Psychology
- Stopping Smoking
- Low Sexual Desire
- Relationships
- Child Development
- Therapy Center NEW
- Diagnosis Dictionary
- Types of Therapy

Understanding what emotional intelligence looks like and the steps needed to improve it could light a path to a more emotionally adept world.
- Coronavirus Disease 2019
- Affective Forecasting
- Neuroscience

An official website of the United States government
The .gov means it’s official. Federal government websites often end in .gov or .mil. Before sharing sensitive information, make sure you’re on a federal government site.
The site is secure. The https:// ensures that you are connecting to the official website and that any information you provide is encrypted and transmitted securely.
- Publications
- Account settings
Preview improvements coming to the PMC website in October 2024. Learn More or Try it out now .
- Advanced Search
- Journal List
- Philos Trans R Soc Lond B Biol Sci
- v.377(1844); February 14, 2022
Experts in action: why we need an embodied social brain hypothesis
Louise barrett.
1 Department of Psychology, University of Lethbridge, Lethbridge, Canada
S. Peter Henzi
Robert a. barton.
2 Department of Anthropology, University of Durham, Durham, UK
Associated Data
This article has no additional data.
The anthropoid primates are known for their intense sociality and large brain size. The idea that these might be causally related has given rise to a large body of work testing the ‘social brain hypothesis'. Here, the emphasis has been placed on the political demands of social life, and the cognitive skills that would enable animals to track the machinations of other minds in metarepresentational ways. It seems to us that this position risks losing touch with the fact that brains primarily evolved to enable the control of action, which in turn leads us to downplay or neglect the importance of the physical body in a material world full of bodies and other objects. As an alternative, we offer a view of primate brain and social evolution that is grounded in the body and action, rather than minds and metarepresentation.
This article is part of the theme issue ‘Systems neuroscience through the lens of evolutionary theory’.
1. A tale of two brains
Today, the social brain hypothesis (SBH) is well established as an explanation for the link between large brains and intense sociality among the anthropoid primates [ 1 – 5 ]. The SBH argues that the need to live in large groups selected for increased brain size and, by extension, the cognitive capacities needed to ensure that these groups remain functional and cohesive. In other words, it is an evolutionary hypothesis that explains how primates have solved the ecological problem of predation risk through the evolution of group-living, [ 6 , 7 ], and then solved the problem of inter-individual competition—which arises inevitably when animals are forced to live in close proximity to one another—by evolving large brains and complex cognitive capacities [ 1 – 5 ]. Support for this hypothesis has come from comparative studies of brain size and social life, in which Robin Dunbar and colleagues [ 1 – 5 ] have played a major role, as well as from studies of primate social behaviour and cognition, in both the wild and captivity [ 8 – 16 ]. In the case of the latter, the evidence presented is not directly tied to brains as such; rather, the objective is to establish the existence of the kinds of cognitive capacities that only a large brain can support (e.g. understanding of third-party relations [ 10 , 11 ], attribution of mental states to others, also known as ‘theory of mind’ [ 14 ] and other forms of perspective taking [ 9 ], tactical deception [ 15 ] and cooperation [ 8 , 12 ]).
Back in the late 1980s and early 1990s, however, another SBH was on offer [ 17 – 19 ]; one that was less concerned with functional explanations for why large brains have evolved in the primate order, and was instead focused on the question of whether regions of the primate brain were specialized for sensing and responding to particular kinds of bodily social stimuli—facial expression, eyes gaze, head and body orientation and biological motion [ 17 – 19 ]. This view of the social brain was associated mainly with the work of Lesley Brothers, along with David Perrett [ 17 – 23 ], where the aim was to establish what particular circuits of the primate brain were doing, and whether these were dedicated to a specific category of objects—other animate beings—as distinct from the broader category of physical objects.
Over time, this latter conception of the social brain has slipped from view in the study of primate social cognition, and Dunbar's more cognitively oriented version of the SBH is what most people now think of when they think of primate brain evolution. This is perhaps understandable given that, as noted above, many studies of primate cognition are aimed at establishing the existence of advanced cognitive capacities that can justify the necessity for large, expensive brains and, in many cases, to identify these as likely pre-cursors of unique human cognitive capacities, like language. To give just one illustrative example, Seyfarth & Cheney [ 24 ] have argued that the origins of spoken language can be found in the structure of baboons' social knowledge, which is construed as a language of thought. Specifically, that, ‘[w]hen a baboon hears vocalizations, she forms a mental representation of call meaning. The meaning of a call sequence includes the representation of an actor who performs a specific action on a recipient and causes the recipient's response. These discrete elements are combined according to the ‘rules’ of call delivery to create a message whose meaning is more than just the sum of the meanings of its constituent elements' [ 24 , p. 7]. This then licenses the conclusion that ‘[a] baboon's assessment of call meaning thus constitutes a discrete, combinatorial, rule-governed and open-ended system of communication in which a finite number of signals can yield a nearly unlimited number of meanings' [ 24 , p. 7], a definition that non-coincidentally maps neatly onto the definition of human language, such that ‘several of the cognitive mechanisms that have long been thought to mark a clear separation between language and non-human primate communication can, in fact, be found—in admittedly simpler form—in the communication and social cognition of non-human primates.’ [ 24 , p. 7]. Thus, although sensitivity to social signals and cues obviously informs the design of these studies, and is also relied on to provide the empirical evidence, the questions of interest are not related to a sensitivity to social signals/cues per se . The focus, instead, is on whether the animals possess the ability to make inferences about the underlying causes that produce these cues, and then to generate further predictions about others' behaviour on the basis of such inferences. Further, no cognitive mechanisms are specifically identified in most studies. Instead, Dennett's intentional stance [ 25 ] is used to inform this methodological strategy, which requires only that animals' behaviour can predicted accurately on the grounds that they behave ‘as if’ they possess the capacity in question (e.g. ‘listeners responded as if they parsed a call sequence as a dramatic narrative’ [ 26 , p. 152] and ‘in their natural behaviour, therefore, non-human primates certainly act as if they are capable of thinking (as it were) in sentences' [ 26 , p. 151] (emphasis added). Cognitive complexity of this order thus supports the SBH by offering evidence for the kinds of social strategizing that allows large social groups to be maintained, and for individual animals to thrive within them.
The current incarnation of the SBH also identifies high-level cognitive abilities (such as mentalizing and the inhibition of prepotent responses) as key to maintaining cohesive social groups, and stresses that these abilities are tied to brain areas unique to primates, specifically the so-called ‘default mode network’ and frontal pole (the most anterior part of the pre-frontal cortex), respectively [ 5 ]. There is also a continued emphasis of the SBH on the neocortex as the comparative neuroanatomical measure that best reflects cognitive ability [ 4 ], which itself reflects the anthropocentric origins of the hypothesis [ 27 ] and the idea that the neocortex is ‘the crowning achievement of evolution and the biological substrate of human mental prowess' [ 28 , p. 274].
Here, we wish to make a case for reincorporating a more Brother's-like view into theories of primate brain evolution for three inter-related reasons: (i) recent comparative analyses have called into question the link between group size and neocortex size in the terms put forward by the SBH [ 29 – 31 ], as well as demonstrating the importance of non-cortical areas, particularly the cerebellum, in primate brain evolution [ 32 , 33 ]; (ii) there is growing recognition that brains evolved first and foremost to control bodies, such that cognition is better conceived of as a set of processes that mediate the adaptive control of bodies in dynamic, unpredictable environments—so-called ‘4E cognition’ [ 34 – 41 ]—and a move away from the traditional ‘disembodied’ view of cognition as a purely brain-based process involving the elaboration, manipulation and transformation of mental representations of the outside world; and (iii) the concept of ‘neural reuse’ [ 42 , 43 ], which suggests that much local neural structure is evolutionarily (and developmentally) conserved, but combined and recombined in different ways across different organisms and species to serve a diverse array of purposes. These three points, therefore, suggest that, rather than looking for human-like cognitive representations in the neocortex of non-humans, a more productive research programme would attempt to understand how both human and non-human cognition emerge from the reuse of systems that have evolved for embodied sensory-motor control.
In what follows, we offer a brief review of recent work on the selection pressures shaping primate brains, consider how work on primate visual and motor systems provides an alternative view of the complexities of primate social life, and conclude with a consideration of some of the implications of non-cortical, particularly cerebellar, evolution. Our suggestion is that a focus on embodied action may open up our understanding of primate brain neurocognitive evolution in ways that recognize the somewhat messy and mosaic nature of the evolutionary process, help identify the relevant differences between apes and other primate species, and enable us to identify more accurately the unique features of hominin cognitive evolution.
2. Multiple pressures, mosaic evolution
Although the relationship between brain size and group size that supports the SBH has been described as ‘remarkably robust’ [ 4 , p. 5], recent comparative analyses do not support such a conclusion. One such study, using a much larger sample of primates (more than 140 species, tripling the samples of previous analyses) and more sophisticated statistical analysis, showed that brain size, after controlling for body size and phylogeny, was best predicted by diet, with frugivores having larger brains than folivores [ 29 ]. By contrast, no measure of sociality (mating system, social system, average group size per species) could be linked to brain size. This outcome is thus more consistent with hypotheses concerning the cognitive demands imposed by extractive foraging of fruits and seeds and the complex spatio-temporal distributions of fruiting trees, than with the cognitive demand of sociality. The study concluded that it was the primary demands on foraging efficiency that then set the stage for selection on social skills [ 29 ]—a conclusion that reverses the SBH argument that animals solve their ecological problems by means of social strategies.
Similarly, using two large comparative datasets, Powell et al . [ 30 ] found evidence that, while brain size was positively associated with home range size, frugivory and activity period, there was no relationship with social group size. More importantly, the authors stressed that all these results were rather unstable and sensitive to the use of different predictor datasets; an outcome they discussed in relation to data quality, intra-specific variance in group size and ecology, statistical power and, crucially, the fact that different selection pressures probably acted on different brain systems at different times. This led them to caution against drawing strong inferences from any attempt to tie brain size evolution to individual selection pressures and to any general measure of cognitive ability. In this regard, it is notable that the original brain size-group size relationship may have been largely dependent on the particular sample of species included in the smaller dataset used in these studies [ 30 ].
This sentiment has been echoed by Wartel et al . [ 31 ] in their analysis of the drivers of brain size, as they also found inconsistent results to be the rule rather than the exception. Given this, they concluded that there is no compelling reason to favour any the adaptive hypotheses on offer—all could either be supported or refuted depending on the datasets used, the choice of variables to include, and the particular observations used to populate those variables [ 31 ]. Although they considered relationships between sensory and motor systems and brain size likely to be robust—for example, a more motor-flexible animal will possess a more complex musculature, with more brain tissue devoted to its control (e.g. [ 44 ])—identifying the drivers of brain size and ‘mental’ capacities are harder to define with ‘non-controversial precision’ ([ 31 ], see also [ 45 ]). These findings also link to the earlier suggestion that ‘cortical ballooning’ (i.e. the higher ratio, or increased proportion, of neocortex to the rest of the brain) as seen in large and large-brained mammals is most parsimoniously understood as being driven by allometric scaling rules that conserve function, rather than by any special selection pressure on cognitive abilities [ 33 ]. Phylogenetically controlled analyses show a strong correlation between body size and the proportion of the brain that is neocortex—that is, large animals possess large neocortices—which seems to reflect a need for more brain space to be devoted to making cortical connections. Larger cortices are made up of more white matter than grey matter, and this white matter consists of fibres making long-range connections that require greater axon diameter and more myelination in order to preserve processing speeds over longer conduction distances [ 33 ]. Ratios between brain structure volumes are confounded by such allometric relationships, and contrary to the underlying assumption of their use, they do not correlate with relative computational capacity as measured by numbers of neurons [ 33 ].
What all these analyses show, then, is that far from neocortex size being ‘a robust predictor of both social group size and many other aspects of social behaviour’ [ 46 , p. 2], the relationship flickers in and out of view, depending on sample size and composition, measures used and choice of predictor variables. This being so, we need to move away from large-scale comparative analyses aimed at identifying a single key driver of overall brain size across the primates, recognize the mosaic nature of primate brain evolution and look instead to the clade-specific ways in which brain size reflects variation in specific neural systems [ 32 , 45 , 47 ].
This is where ideas relating to neural reuse become immediately relevant. If we look at cortico-cerebellar networks, for example, we see reciprocal loops that have a remarkably similar anatomical architecture across multiple systems, and which appear to have evolved by duplicating circuits and retaining a powerful, general functional organization. In particular, these cortico-cerebellar loops mediate forward models, whereby the system makes predictions about the consequences of actions and continuously adaptively adjusts behaviour in real time [ 48 ]. Originally developed within the framework of control theory and applied to understanding sensory-motor control, it is now understood that forward models present a solution to the problem of behavioural control more generally. The precise function of a cortico-cerebellar loop thus depends on its specific connections (e.g. with motor versus pre-frontal cortex). Hence, the cerebellum is now known to be involved in a wide range of functions, including motor control, perception, language, working memory, cognitive control and social cognition [ 49 ]. This dissolves the arbitrary distinction between cortex as the ‘thinking’ part of the brain and the cerebellum as engaged only in sensorimotor control, and allows us to consider the range of tasks in which the cerebellum is involved, rather than worrying about whether or not the cerebellum is ‘cognitive’ in the manner of the neocortex [ 32 , 33 ].
In terms of primate brain evolution, the coordinated functioning of neocortex and cerebellum is reflected in a strong pattern of correlated evolution between them, independent of change in other regions [ 33 ]. Although it is often said that an expanded neocortex is what marks humans out evolutionarily, comparative analysis reveals more rapid cerebellar relative to neocortical expansion in human and great ape lineages than in other primates [ 32 , 33 ]. Cerebellar expansion in apes is reflected in their slow maturation, and cerebellar and neocortical size display distinct life-history correlates congruent with their developmental schedules [ 50 ]. In particular, cerebellum shows relatively substantial post-natal growth, particularly during the juvenile period, and its evolutionary expansion in apes corresponds to significantly slower post-natal development [ 50 ]. This suggests that environmental input is crucial for cerebellar function, pointing to the importance of well-coordinated action in the world as a key feature of primate social life, and a probable selection pressure on brain evolution. We return to the topic of the cerebellum below in our consideration of human uniqueness.
3. Social coordination in physical space
Although the general relationship between neocortex size and group size has not withstood more extensive and rigorous analyses, the relationships detected between brain size, frugivory and activity period are in line with earlier work suggesting that visual specialization has been one of the important drivers of primate brain size. For example, independent correlations between relative neocortex size, activity period (diurnal versus nocturnal), degree of frugivory and group size [ 51 ] have been shown to reflect the relative expansion of the geniculo-cortical visual system and enhancements of fine-grained binocular vision [ 52 , 53 ].
These analyses found that both V1 and non-V1 cortex were positively correlated with encephalization [ 54 ], but only in association with the parvocellular layers of the lateral geniculate nucleus (LGN), which is the pathway mediating vision for fine detail and colour, not the magnocellular layers involved in movement detection. The relative number of neurons in the parvocellular layers of the LGN was also found to correlate with social group size, as well as with diurnality and degree of frugivory, whereas the magnocellular layers did not. Thus, there is evidence for increasing visual specialization that was linked to both social and ecological selection pressures. This accords with the proposal that visual processing of complex and rapid social interactions, and the monitoring of visual signals (most prominently facial expression and eye-gaze) were the skills linked to social group size maintenance [ 33 ]—a suggestion in line with earlier, more detailed work on specific regions of the brain [ 17 – 23 ]—while at the same time underpinning the fine visual discrimination and coordination needed for manipulating small fruits and seeds. That is, certain aspects of primate brain size evolution can be tied to the socioecological demands of the perceptual domain quite directly, rather than being a response to a pressing need for more ‘abstract’ sociocognitive skills: a ‘visual brain hypothesis', if you will. It should also be noted that as variation in neocortex size is also strongly related to the evolution of the cerebellum as well as visual structures, it is perhaps more accurate to describe this as a visuo-motor brain hypothesis [ 33 ].
With respect to a reconceptualization of the SBH along more embodied lines, we suggest that the findings above steer us toward a ‘second person participatory’ perspective on social interaction [ 54 – 56 ], i.e. one that concerns itself with how animals interact with other physically, rather than with the generation of inferences about the inferences they might generate about each other. Here, cognition is construed as flexible action in the world, not as internal propositions directed at the world. Consequently, neurophysiological and behavioural work on motor coordination, and in particular, peripersonal space (PPS) [ 57 – 59 ] also offers a promising avenue of exploration with respect to understanding the demands of social coordination in the physical environment. This is work that can be married to the evidence on visual specialization that emphasizes the importance of monitoring facial expression, eye-gaze and other social signals.
The term ‘peripersonal space’ (or ‘near space’) was first used to describe the area within an arm's reach of the body. In this original conception, PPS designated the range within which physical objects could be grasped and manipulated [ 60 ]. As work has continued, however, this notion of PPS has been expanded and refined, as we describe below. To characterize PPS more precisely in neurobiological terms, studies on monkeys have identified neuronal populations in parietal cortex, including the ventral intraparietal area (VIP) and area 7b, areas of frontal cortex, including ventral premotor cortex (vPMC) and areas 6 and F4, as well as subcortical areas, like putamen [ 58 ]. These are areas that respond not only to tactile (somatosensory) stimuli, but also to visual or auditory stimuli presented in close proximity to the somatosensory receptive field (it is worth noting here that the bimodal nature of these neuronal responses undermines the notion that perception and action can be seen as separable processes). These receptive fields are ‘anchored’ to a specific body part (e.g. the face, the hand, the arm), forming a body-part-centred ‘mapping’ of space around the body (see [ 58 , 59 ] for review). The responses of these multi-sensory neurons, and the notion of PPS, was thus argued to be crucial for guiding effector-specific movements in relation to nearby objects [ 59 – 63 ]. Further work, however, found evidence for a defensive function of PPS—where, for example, sudden puffs of air presented near the face of macaques produced distinctive startle responses that could also be produced by stimulation of the neurons in the VIP, and the polysensory zone [ 64 , 65 ]. Thus, in addition to guiding action with respect to non-threatening objects, PPS is argued to provide a ‘margin of safety’ that allows animals to avoid threatening objects (including conspecifics) as well as collisions with physical hazards [ 66 ].
Much of the early work on PPS depicted it as a series of clearly demarcated ‘bubbles’ extending from particular areas of the body—something suggested to be a legacy of the very earliest work on ‘flight zones' in animals [ 67 ] and work on proxemics and the notion of ‘personal space’ in humans [ 68 ]. However, more recent work has shown that most multi-sensory neurons show a graded, continuous response with respect to distance from the body, rather than a simple stepwise ‘in-or-out’ response [ 57 ], while some cells in the vPMC have receptive fields that extend further than reaching distance (sometimes even to the end of the testing room) [ 57 ]. PPS does not seem to be confined to the narrow zone within arm's reach of the body (which was an artefact of design of the earliest studies), but shows a certain degree of flexibility, where PPS-related neurons not only respond to stimuli with graded (or even reversed) relationships to distance, but are also influenced by factors such as the motion of body parts, and the trajectory and valence of the stimulus [ 57 ]. As such, PPS is better conceived as a ‘set of continuous relevance-estimation fields', or zones in space that reflect the behavioural relevance of actions aimed at either making contact or avoiding contact between objects (including conspecifics and other animals) and the body [ 57 ], and where the size and extent of such fields is adjusted continuously in relation to ongoing activity. Classic work on macaques, for example, showed that, following training with a rake-like tool that allowed them to retrieve distant food, the receptive fields of the visual neurons associated with the somatosensory receptive field of the hand were extended to include the length of the tool [ 69 , 70 ].
Work on PPS in general, and the field-theory of PPS in particular [ 57 ], offers us the kind of embodied view mentioned above in §1. More specifically, it pushes us away from the standard view of behaviour as a stepwise process that proceeds linearly from sensory input to motor output, and towards the kind of embodied, interactive approach described by affordance competition theory [ 71 ]. Here, the neural architecture that mediates behaviour is best characterized as a set of competing sensorimotor loops, with multiple simultaneous competing actions constantly being prepared, and selection among them occurring in response to the possibilities for action (affordances) in the environment, and how available affordances shift as part of the ongoing flow of activity [ 71 ]. Bufacchi & Iannetti [ 57 ] use a human example to illustrate this, in which they consider how the PPS field derived from reaction times to a tactile stimulus on the hand (i.e. pressing a button as soon as the stimulus is felt) varies as an auditory stimulus approaches, with reaction times being faster when the sound is closer. As they note, it is not immediately obvious why pressing a button should have anything to do with creating or avoiding contact between the hand and the sound source. However, if multiple simultaneous competing actions are being prepared at all times then, as sounds move closer to the hand, actions related to making or avoiding contact become increasingly relevant. Given that similar actions will share similar neural network activity, then any actions produced by tactile stimulation (here, a button press) should share its network with actions that create or avoid contact with the sound source. As a result, as the sound approaches, any hand-related actions should be more readily enacted, giving rise to shorter reaction times. Their argument, then, is that most PPS fields will result from the summation of the relevance of a whole set of possible actions, rather than being linked to a single specific action.
Hunley & Lourenco [ 59 ] similarly consider PPS to be a body-centred network responsible for the coordination of actions that both enables interactions with, and avoidance of, objects and other living entities. In their view, this network is described by distinct defensive and non-defensive pathways that, although dissociable, nevertheless display evidence of shared organization, and can be viewed as part of a single integrated system. In this regard, they can be considered as somewhat akin to dorsal and ventral pathways in the visual system [ 72 , 73 ]. Evidence from monkeys indicates that non-defensive behaviours are associated with a pathway that includes the anterior parietal area (AIP), area 7b, and premotor area F5, while defensive behaviours are associated with a pathway involving VIP and F4 [ 74 ]. There is also evidence that these pathways may correspond to different body parts, with AIP, 7b and F5 largely tied to activity related to the hand and arm, while VIP and F4 are linked to the head and face: activities that involve interactions towards and with objects and others, and those that involve protecting vulnerable areas of the body, respectively [ 74 ]. The defensive and non-defensive pathways can be conceptualized similarly to a dimensional view of neophilia and neophobia: although these are often seen as the extreme ends of a single continuum, they are better viewed as two separate dimensions, because animals require some combination of both traits to function effectively [ 75 ]. Just as an animal may need to be neophilic enough to exploit novel situations but also be sufficiently neophobic to do so in ways that protect them from danger, so PPS represents two dimensions that allow animals both to navigate the world safely, while simultaneously providing the capacity to effectively manipulate and exploit physical and social resources in adaptive and functional ways.
In our view, then, work on PPS provides a neurobiologically grounded solution to the problem of coordination through physical and social space that, in current conceptions of the SBH, relies on executive processes like inhibition and inferential reasoning. It should be readily apparent that a flexible sensitivity to the area of space around the body, and the selection of relevant actions, is crucial to the various demands of primate social life in the round: the selection and processing of particular foods, movement in and through the trees, and the ongoing back and forth of social interactions, whether this be monitoring and withdrawing from threatening encounters or coordinating affiliative behaviours like grooming, mating and infant care. One can readily see how visual specializations linked to detecting gaze direction and facial expression would form part of a coordinated sensorimotor system that allows animals to regulate their behaviour with respect to the approach and avoidance of their conspecifics. Shifts in the extent of both defensive and non-defensive PPS would thus enable the specification and selection of appropriate actions as the animal moves through space.
Our argument here thus resembles Cisek & Pastor-Bernier's [ 76 ] critique of neuroeconomics. The latter is a field where a great deal of energy is spent debating whether modern human decision-making is ‘economically rational’ and determining the kinds of deliberative cognitive mechanisms that would be required to calculate values and commit to rational action. Much of this work is focused on executive functions considered to take place in the frontal lobes and separate from sensorimotor control; reasoning that is not unlike that used in the SBH, which as we have noted, places a similar emphasis on high-level executive processes. Such a stance seemingly makes the implicit assumption that brain evolution is a process by which a ‘mammalian’ cerebral cortex has been draped over a more primitive ‘reptilian’ brain, even though this view has long since been debunked [ 77 ]. Evolution does not proceed by bolting new structures onto those already existing, but by the elaboration and specialization of ancestral neurological circuits, in ways that preserve the system's overall integrity [ 76 ]. The theory of neural reuse can also be folded into this, again offering an alternative neurobiological picture compared to the SBH. A true evolutionary perspective, then, pushes us to build theories that are addressed to the kinds of ‘embodied decisions’, captured by the examples described above: decisions that occur during ongoing activity, and which are mediated by sensorimotor processes that enable close coordination with the environment.
Consider a group of baboons at the beginning of the day: some remain on the sleeping cliffs grooming, others are already foraging on the grassy plain below, a few are beginning to drift off on the day's journey. Now, consider a baboon that encounters this social scene: there are decisions to make concerning hand and foot placement as she descends from the cliff while, at the same time, there are decisions about which animals she can safely approach along the route, and those to whom she should give a wide berth. Once on the ground, there are decisions to make about where to forage—which means whether and where to dig for corms—and this means monitoring who else is around, and where, and what they are doing and with whom. All this might mean having to decide where to move as others approach or move further away, and all the while she is controlling and coordinating her hand movements, as she digs and plucks corms from the ground, dusts them off on her arm, peels away tough outer layers with fingers and teeth. This baboon is constantly in action and in the moment, responding in real time to a perpetual flow of socio-environmental stimuli.
These kinds of embodied decisions are different from those considered in the highly cognitive, anthropocentric view of neuroeconomics [ 76 ] and, we would argue, the SBH. The options available to the baboon are potential action possibilities—affordances—not abstract ‘values’. Options of this nature are strongly influenced by biomechanical constraints and environmental layouts, and so cannot be considered as clean-cut abstractions occurring in some independent cognitive or mental realm. In addition, decision-making under such conditions is simultaneous: our baboon may encounter a series of objects and social others, but there are always several action possibilities available for selection at any given point. Thus, embodied decisions are always highly dynamic: ‘as an animal moves through its world, available actions are constantly changing, some are vanishing while others appear, and also the relevant variables (outcome values, success probability, action cost) are always in flux…each embodied decision is a single-trial situation with unique settings.’ [ 76 , p. 3]. Our baboon continuously picks up sensory information about relevant affordances as part of her ongoing activity, with no temporal distinction between choice and implementation—in many ways, the choice is the implementation. In other words, decisions need not be determined by a central executive of any kind, but depend instead on which of the reciprocally connected sensorimotor networks are the first to ‘commit’ to a given action strongly enough to pull the rest of the network into a ‘distributed consensus' [ 76 ]. The notion of specialized but flexible PPS fields can be folded easily into this process of embodied decision-making: sensitivity to the proximity of threatening and non-threatening objects and others is key to action specification and selection [ 58 ]. In terms of primate brain evolution, then, our proposition is that, as with the analyses of visual specialization [ 51 – 53 ], there is much merit to be had in analyses that investigate the size and structure of parietal regions associated with PPS in relation to the size and structure of social groups (e.g. in relation to strength of dominance hierarchies, uni-male versus multi-male, multi-female structures, single-level versus multi-level societies), as well as the demands of the foraging environment, and the degree of terrestriality. We also need to investigate and understand potential links between the visual brain and PPS-related regions of the brain.
4. Emergent systems of human social cognition
In addition to offering new avenues of exploration with respect to primate brain evolution, thinking of the social brain as an integrated system of sensorimotor coordination can offer new insights into the evolution of our own cognition. Graziano [ 61 ], for example, emphasizes the importance of PPS and physical coordination as the basic grounding for more sophisticated forms of human social cognition: ‘personal space plays its most profound human role in the social domain. That invisible bubble of protected space, the space in which you do not want other people, creates the scaffold for all other social interactions. It places us in a great social honeycomb of decorous relationships' [ 61 , p. 147]. Experimental data on humans tends to support Graziano's argument. A study using a cross-modal visual–tactile task to determine the bounds of PPS found that PPS expanded following an interaction with another person, but only if the action was cooperative—in other words, low-level sensorimotor processing modulated processes of high-level social cognition [ 78 ]. As a non-threatening interaction, one might imagine that cooperation would shrink PPS fields around the body, not extend them. However, we need to consider defensive and non-defensive pathways as connected but distinct [ 59 ]. While cooperative interactions should indeed involve a reduction in defensive PPS, non-defensive PPS should expand to better facilitate social coordination in the context of cooperation. Thus, the other person plays a functional role within PPS, in much the same way that monkey PPS was shown to extend to include raking tools. There is also evidence that PPS can vary in relation to particular psychological traits: expansion of PPS following tool use was affected by expressed levels of claustrophobic fear, with participants that scored high on this trait experiencing less expansion [ 79 ].
De Jaegher [ 80 ] offers similar arguments in her enactive account of autism, which posits that sensory and motor difficulties are basic to autism, rather than seeing them as separate from deficits in so-called ‘higher cognitive functions', such as theory of mind. Specifically, she suggests that ‘sensorimotor interactional coordination ability’ lies at the base of our ability to engage in ‘participatory sense-making’, defined as the way that neurotypical people make sense of the world by moving around in it and with it. How, in other words, we coordinate our movements with others when interacting, such that we generate and transform meaning together. Differences in how one perceives and moves through the world may, therefore, lead to difficulties in coordinating social interactions, and hence to failures in the participatory sense-making needed to understand ourselves and others. Among other things, De Jaegher [ 80 ] discusses how, in comparison to typically developing children and those with Aspergers, autistic children display lower levels of visuopostural attunement to the environment [ 81 , 82 ]. Other researchers have also suggested that motor problems are basic to autism: children with autism experience difficulties with ‘reach-to-grasp’ movements, showing both poorer planning and execution [ 83 ]. These kinds of visuo-motor deficits are suggested to be related to anomalies in magnocellular pathways, visuo-cerebellar circuits, as well as in cerebellar–premotor cortex loops [ 83 ].
More recent work has specifically considered PPS in the context of autism. For example, using an audio-tactile integration task to assess PPS, adults with autism spectrum disorders (ASDs) were found to have smaller PPS than non-ASD adults, and one with a sharper boundary [ 84 ]. This was also associated with a lower susceptibility to the full body illusion [ 84 ]. In a comparison of autism and schizophrenia as disorders of the self, and how this might relate to PPS, it was argued that these conditions might sit at either end of a continuum, with autism reflecting a steep self-other gradient, with little variation, while schizophrenia reflects a shallow self-other gradient, associated with heightened variability in PPS [ 85 ] (schizophrenic individuals, for example, are more susceptible to the rubber hand illusion [ 86 , 87 ]). Such work, therefore, supports the argument that human cognitive capacities emerge from the elaboration and specialization of ancestral neurobiological circuits that enable the control of bodily actions in the environment. The deficits seen in autism and schizophrenia point to fundamental issues in coordinating physical and social space, which in turn are tied to issues relating to the nature of the self, and the recognition and maintenance of self-other boundaries [ 85 , 88 ]. In such a view, these conditions are not wholly psychological in nature, as this is usually understood, i.e. distinct from physical problems in the body. Rather they demonstrate how particular psychological conditions cannot be separated from how people monitor and move physically in space.
5. Cerebellar expansion, neural reuse and human uniqueness
In conclusion, we return to the role of the cerebellum in neurocognitive evolution and outline the implications for understanding what appear to be specialized human capacities. Given the rapid cerebellar expansion during ape evolution, the role of the cerebellum in organizing and comprehending action sequences [ 89 ] and apes' marked facility for sequence organization in below-branch brachiating locomotion, tool use and extractive foraging [ 89 ], it seems reasonable to propose that there has been selection for greater temporal organization, comprehension and learning of behavioural sequences in these taxa. This suggests that, in addition to placing greater emphasis on sensorimotor coordination in studies of primate brain evolution in general, we also need to give more serious consideration to ideas relating to ‘technical intelligence’ when considering ape and human brain evolution in particular. This includes the idea that the syntactical features of language may have been built from pre-existing sensorimotor specializations common to all apes [ 90 ], where both social and technical skills are important, and no doubt interact with each other in mutually reinforcing ways [ 33 ]. The neural reuse perspective can easily accommodate such a scenario: cortico-cerebellar loops involved in organizing coherent sequences of movements to produce tools could be repurposed to organize coherent sequences of social actions to produce social outcomes, or coherent sequences of words to produce sentences
Indeed, Ellis [ 91 ] offers precisely this argument, proposing an evolutionary scenario that links our remarkable manual dexterity and unparalleled ability to manipulate objects within our PPS (which he refers to as ‘toolspace’) to the construction and use of tools and other artefacts of various kinds. Specifically, physically coordinated, joint activity on objects is argued to have given rise to forms of social communication that eventually became symbolic language. Consequently, in a reversal of the standard view, language and other complex cognitive skills do not form the underpinnings of our sophisticated material cultures but are, instead, considered to be their manufactured products [ 91 ]. Human cognition is, therefore, ‘in large part a cultural artefact invented by successive generations of especially dextrous primates' [ 91 , p. 1]. As with our argument above, the emphasis here is on sensorimotor coordination as the foundation of, and scaffold for, our characteristically human cognitive skills. More broadly, this theory again resonates with ideas of neural reuse, where neural adaptations build upon one another and specific adaptations have multiple effects on behaviours traditionally considered to represent different ‘domains’. That is, it seems possible to build on, or extend from, Ellis' theory [ 91 ], and investigate more closely how cortico-cerebellar networks potentially may support similar types of processes in different contexts, such as sequencing and prediction in brachiation, tool making and language.
Indeed, complex sequence organization and learning has long been proposed as a key aspect of human cognition that underlies tool use, language, music and other distinctive human capabilities [ 92 ]. More recently, Ghirlanda et al . [ 93 ] found evidence that the ability to learn arbitrary stimulus sequences distinguishes humans from non-human species. Unlike humans, where the correct representation and processing of sequential information is crucial to all kinds of everyday tasks, and also to some of our more rarefied skills, such as the creation of music and mathematics, non-human animals do not seem to be able to learn and make use of arbitrarily sequential information. For example, in studies comparing humans and macaques on the same task, humans took 30 trials per presented sequence to achieve 90% correct responses, whereas macaques required on the order of 400 trials per sequence to achieve 70% correct. Notably, however, there are no studies, to date, investigating these abilities in non-human apes. If the findings on cortico-cerebellar evolution are pointing us in the right direction, however, we might predict that apes will show improved skills in this domain.
In addition, although their survey of empirical findings, combined with a model of non-human sequence discrimination, provided good evidence to indicate that humans show a greatly improved memory for sequence formation, Ghirlanda et al . [ 93 ] did not show how humans have overcome the limitations of non-human memory. One suggestion that they offer is that language provides the kind of explicit verbal scaffolding needed to deal with arbitrary sequences (e.g. ‘respond when you see blue first, and then yellow, but not if this comes before red’). However, as they also point out, this puts the cart before the horse, as language itself is dependent on sequence formation, and such verbal strategies could not work without understanding of concepts such as ‘first’ and ‘before’. What they do not consider, however, are the kinds of ‘external’ strategies suggested by Ellis, in which the processing and making of material artefacts, and the capacity for joint action, might have scaffolded these skills, not least by generating forms of material culture that physically embody such sequences, and thereby provide a form of external memory (see [ 94 ] for an explanation of just such a process in the invention of number concepts). It may be that we are only capable of particular kinds of representational thinking (e.g. thinking in terms of the intersection in a Venn diagram) because we first generated such things in physical form, and were then able to internalize them (whatever ‘internalize’ might mean). One sees something similar in the counting practices shown by children who learn arithmetic using an abacus, and can then perform remarkable feats of ‘mental arithmetic’ without the abacus present, including complex multiplication and long-division. What is most notable here, perhaps, is how the children continue to perform the characteristic hand movements corresponding to moving beads along the bars of the physical abacus as they engage in these ‘mental’ calculations [ 95 , 96 ].
In summary, our argument, then, is that characteristic and complex forms of human cognition may ultimately depend on basic processes of physical coordination in relation to objects and social others. Investigations of primate brain evolution will, therefore, benefit from focusing more closely on how the brain has become specialized for socially and physically oriented forms of sensorimotor coordination, and how these may have scaffolded and set the stage for the evolution of unique human cognitive capacities.
Data accessibility
Authors' contributions.
All authors gave final approval for publication and agreed to be held accountable for the work performed therein.
Competing interests
We declare we have no competing interests.
Funding for this study came from NSERC Discovery Grants awarded to L.B. and S.P.H., and a Canada Research Chair (Tier I) award to L.B.
- Share full article
Advertisement
Supported by
Working With Your Hands Is Good for Your Brain
Activities like writing, gardening and knitting can improve your cognition and mood. Tapping, typing and scrolling? Less so.

By Markham Heid
The human hand is a marvel of nature. No other creature on Earth, not even our closest primate relatives , has hands structured quite like ours, capable of such precise grasping and manipulation.
But we’re doing less intricate hands-on work than we used to. A lot of modern life involves simple movements, such as tapping screens and pushing buttons, and some experts believe our shift away from more complex hand activities could have consequences for how we think and feel.
“When you look at the brain’s real estate — how it’s divided up, and where its resources are invested — a huge portion of it is devoted to movement, and especially to voluntary movement of the hands,” said Kelly Lambert, a professor of behavioral neuroscience at the University of Richmond in Virginia.
Dr. Lambert, who studies effort-based rewards, said that she is interested in “the connection between the effort we put into something and the reward we get from it” and that she believes working with our hands might be uniquely gratifying.
In some of her research on animals , Dr. Lambert and her colleagues found that rats that used their paws to dig up food had healthier stress hormone profiles and were better at problem solving compared with rats that were given food without having to dig.
She sees some similarities in studies on people, which have found that a whole range of hands-on activities — such as knitting , gardening and coloring — are associated with cognitive and emotional benefits, including improvements in memory and attention, as well as reductions in anxiety and depression symptoms.
These studies haven’t determined that hand involvement, specifically, deserves the credit. The researchers who looked at coloring, for example, speculated that it might promote mindfulness, which could be beneficial for mental health. Those who have studied knitting said something similar. “The rhythm and repetition of knitting a familiar or established pattern was calming, like meditation,” said Catherine Backman, a professor emeritus of occupational therapy at the University of British Columbia in Canada who has examined the link between knitting and well-being.
However, Dr. Backman said the idea that working with one’s hands could benefit a person’s mind and wellness seems plausible. Hands-on tasks that fully engage our attention — and even mildly challenge us — can support learning, she added.
Dr. Lambert has another hypothesis. “With depression, people experience something called learned helplessness, where they feel like it doesn’t matter what they do, nothing ever works,” she said. She believes that working with one’s hands is stimulating to the brain, and that it could even help counteract this learned helplessness. “When you put in effort and can see the product of that, like a scarf you knitted, I think that builds up a sense of accomplishment and control over your world,” she said.
Some researchers have zeroed in on the possible repercussions of replacing relatively complicated hand tasks with more basic ones.
In a small study of university students published in January, Norwegian researchers compared the neurological effects of writing by hand with typing on a keyboard. Handwriting was associated with “far more elaborate” brain activity than keyboard writing, the researchers found.
“With handwriting, you have to form these intricate letters by making finely controlled hand and finger movements,” said Audrey van der Meer, one of the authors of that study and a professor of psychology at the Norwegian University of Science and Technology. Each letter is different, she explained, and requires a different hand action.
Dr. Van der Meer said that the act of forming a letter activates distinctive memories and brain pathways tied to what that letter represents (such as the sound it makes and the words that include it). “But when you type, every letter is produced by the same very simple finger movement, and as a result you use your whole brain much less than when writing by hand,” she added.
Dr. Van der Meer’s study is the latest in a series of research efforts in which she and her colleagues have found that writing and drawing seem to engage and exercise the brain more than typing on a keyboard. “Skills involving fine motor control of the hands are excellent training and superstimulation for the brain,” she said. “The brain is like a muscle, and if we continue to take away these complex movements from our daily lives — especially fine motor movements — I think that muscle will weaken.” While more research is needed, Dr. Van der Meer posits that understimulation of the brain could ultimately lead to deficits in attention, memory formation and problem solving.
But as with knitting and coloring, some experts question the underlying mechanisms at play.
“With some of this research, I think it’s hard to dissociate whether it’s the physical movement of the hands that’s producing a benefit, or whether it’s the concentration or novelty or cognitive challenge involved,” said Rusty Gage, a professor at the Salk Institute for Biological Studies in San Diego.
Dr. Gage studies how certain activities can stimulate the growth of new cells in the brain. “I think if you’re doing complex work that involves making decisions and planning, that may matter more than whether you’re using your hands,” he said.
That said, the benefits of many hands-on activities aren’t in doubt. Along with gardening and handicrafts, research has found that pursuits like making art and playing a musical instrument also seem to do us some good.
“You know, we evolved in a three-dimensional world, and we evolved to interact with that world through our hands,” Dr. Lambert said. “I think there are a lot of reasons why working with our hands may be prosperous for our brains.”
IMAGES
VIDEO
COMMENTS
Hence, the relationship between brain size and group size is indirect, and the real functional relationship in the social brain hypothesis is that between brain (or brain region) size and/or wiring and social cognitive abilities or competences that allow primates to manage relationships (Dunbar, 1998, 2011a, 2012a). In effect, group size is an ...
Here we present a theory, captured in an analytic and computational model, that explains these increases in brain size: The Cultural Brain Hypothesis. The theory relies on the idea that brains expand to store and manage more information. Brains expand in response to the availability of information and calories.
This chapter defends a version of the social brain hypothesis, focusing especially on the cognitive powers involved in the 'Theory of Mind' that. ... A primary goal of evolutionary psychology is to understand the evolution of the human mind, our unique cognitive adaptations, and how these translate into cognitive functioning and, ultimately ...
The social brain hypothesis offers an explanation; substantial neocortex volume favored group formation, social complexity, and coalition formation. In fact, Brewer and Caporael (2006) theorize that the group context was essential for human evolution. At the same time, group formation breeds intergroup conflict, and disproportionate cortical ...
The social-brain hypothesis refers to a quantitative relationship between social-group size and neocortex volume monkeys and apes. This relationship predicts a group size of approximately 150 for humans, which turns out the typical size of both social communities in small-scale societies and personal social networks in the modern.
CONCLUSION. The study of brain evolution could benefit greatly from methods and techniques used by cognitive neuroscience and neuropsychology, and in turn, neuropsychology could benefit from incorporating comparative data on human brain evolution. The Inferential Brain Hypothesis is an example of how understanding changes in structure-function ...
The Social Brain. The social brain hypothesis attempts to explain the extraordinary size and complexity of the human brain by appeal to particular pressures that a species adapted to social interaction would have had to face, ranging from deception to cooperation to ways of obtaining food and ensuring offspring (Allman 1999; Barrett & Henzi ...
The social brain hypothesis ... Robin Dunbar is Professor of Evolutionary Psychology and Behavioural Ecology at the University of Liverpool, England. His research primarily focuses on the behavioral ecology of ungulates and human and nonhuman primates, and on the cognitive mechanisms and brain components that underpin the decisions that animals ...
The development of the social brain paradigm reflects a general development from hierarchical to network thinking across the intellectual spectrum during the latter part of the twentieth century. I discuss the evolution of the social intelligence hypothesis into the social brain hypothesis, and the reigning myths about the brain that have ...
This point references Dr. Robin Dunbar's famous Social Brain Hypothesis, where the term "social brain" is used to associate brain size with social complexity in primates and signifies the ...
4. In defence of the interactive brain hypothesis (E.D.P. and H.D.J.) Let us consider the IBH at its most radical: the claim that the dynamics of social interaction play constitutive roles in social cognition. The developmental version seems less controversial, although its implications are not trivial (see e.g. ). In fact, for any ...
Humans, like most mammals, are intensely social. In many ways, primates' success from an evolutionary perspective is a direct consequence of that sociality. Primate societies are implicit social contracts that allow some of the problems of survival and reproduction to be solved co-operatively. Social contracts of this kind work because they allow relevant problems to be solved more efficiently ...
More simply put, the brain can either revise its predictions to match the world or change the world to make the predictions come true. Fig. 1: Beliefs, desires, predictions and precision. a Left ...
This volume brings together two powerful approaches — the social brain hypothesis and the concept of the distributed mind. The volume compares perspectives on these two approaches from a range of disciplines, including archaeology, psychology, philosophy, sociology and the cognitive and evolutionary sciences.
The social brain hypothesis was proposed as an explanation for the fact that primates have unusually large brains for body size compared to all other vertebrates: Primates evolved large brains to manage their unusually complex social systems. Although this proposal has been generalized to all verteb …
Thus was born the "social brain hypothesis" and ... Science articles cover neuroscience, psychology, AI, robotics, neurology, brain cancer, mental health, machine learning, autism, Parkinson's, Alzheimer's, brain research, depression and other sciences. 2 Comments. Luiz Veiga says: September 29, 2021 at 7:49 am. Interesting. gizella szabo says:
Architectures in neural networks commonly assume that inference is hierarchical. In this Perspective, Suzuki et al. present the shallow brain hypothesis, a neural processing mechanism based ...
A primary goal of evolutionary psychology is to understand the evolution of the human mind, our unique cognitive adaptations, and how these translate into cognitive functioning and, ultimately, behaviour. ... One widely cited example of evidence for the social brain hypothesis is that primate social group size correlates with species brain size ...
The hypothesis states that interactive experience and skills play enabling roles in both the development and current function of social brain mechanisms, even in cases where social understanding happens in the absence of immediate interaction. We examine the plausibility of this hypothesis against developmental and neurobiological evidence and ...
Origins. This field of study has its historical roots in numerous disciplines including machine learning, experimental psychology and Bayesian statistics.As early as the 1860s, with the work of Hermann Helmholtz in experimental psychology, the brain's ability to extract perceptual information from sensory data was modeled in terms of probabilistic estimation.
The imprinted brain hypothesis is an unsubstantiated hypothesis in evolutionary psychology regarding the causes of autism spectrum and schizophrenia spectrum disorders, first presented by Bernard Crespi and Christopher Badcock in 2008. It claims that certain autistic and schizotypal traits are opposites, and that this implies the etiology of the two conditions must be at odds.
The Resistance Hypothesis. ... Ph.D., is an Associate Teaching Professor at Johns Hopkins University in the Department of Psychological and Brain Sciences. He teaches courses and conducts research ...
Syndromic autism refers to autism spectrum disorder diagnosed in the context of a known genetic syndrome. The specific manifestations of any one of these syndromic autisms are related to a clinically defined genetic syndrome that can be traced to certain genes and variants, genetic deletions, or duplications at the chromosome level. The genetic mutations or defects in single genes associated ...
Abstract. A recent framework inspired by phenomenological philosophy, dynamical systems theory, embodied cognition and robotics has proposed the interactive brain hypothesis (IBH). Whereas mainstream social neuroscience views social cognition as arising solely from events in the brain, the IBH argues that social cognition requires, in addition ...
1. A tale of two brains. Today, the social brain hypothesis (SBH) is well established as an explanation for the link between large brains and intense sociality among the anthropoid primates [1-5].The SBH argues that the need to live in large groups selected for increased brain size and, by extension, the cognitive capacities needed to ensure that these groups remain functional and cohesive.
Dr. Van der Meer's study is the latest in a series of research efforts in which she and her colleagues have found that writing and drawing seem to engage and exercise the brain more than typing ...